PROJECT
DESCRIPTION
RESULTS OF PRIOR NSF SUPPORT
Harold P. Batchelder: GLOBEC: Effects of Seasonal and
Interannual Variability on Zooplankton Populations in the
Batchelder, H. P., C. A. Edwards, and
T. M. Powell. 2002.
Individual-based models of copepod populations in coastal upwelling
regions: implications of physiologically and environmentally influenced diel
vertical migration on demographic success and nearshore retention. Prog. Oceanogr., 53, 307-333.
Batchelder, H. P., and T. M. Powell.
2002. Physical and Biological Conditions and Processes in the
Batchelder,
H.P. Forward-in-Time and Backward-in-Time Trajectory (FITT/BITT) modeling of
particles and organisms in the coastal ocean. Submitted to Jour. of Atmos. and Oceanic
Tech.
Perry,
R. I., H. P. Batchelder, D. L. Mackas, S. Chiba, E. Durbin, W. Greve and H. M.
Verheye. 2004. Identifying global synchronies in marine
zooplankton populations: issues and
opportunities. ICES J. Mar. Sci.,
61, 445-456.
James J. Bisagni: (JJB-1): GLOBEC: Satellite-Derived
Estimates of Mixing Across Sea Surface Temperature Fronts in the Georges Bank
Region, (Principal Investigator), NSF/NOAA US
Bisagni,
J. J. and K. W. Seemann, 1999. Visualizing annual and interannual sea surface
temperature variability over the Gulf of Maine/Georges Bank region. Eos, 80:OS264
Mavor,
T. P. and J. J. Bisagni, 1999. Seasonal variability of satellite-derived sea
surface temperature fronts in the vicinity of
Bisagni,
J. J., K. W. Seemann, and T. P. Mavor. 2001. High-resolution satellite-derived
sea surface temperature variability over the
Mavor, T. P. and J. J. Bisagni, 2001. Seasonal variability of sea
surface temperature fronts on
JJB-2: Collaborative
Research: GLOBEC-01:Patterns of energy flow and utilization on
Bisagni, J. J., 2003. The seasonal cycles of nitrate supply and
potential new production in the
Steele,
J.H. and J.S. and Collie 2003. Functional diversity and stability of coastal ecosystems.
In: The Sea, Vol. 13. A.R. Robinson
and K.R. Brink (eds.). In press.
JJB-3: GLOBEC-01:
The physics of
Wishner, K. F., D. J. Gifford, B. K. Sullivan, J. J.
Bisagni, D. M. Outram and
Bisagni, J. J., 2004. Seasonal and interannual variability of the shelf
water-slope water front between 75° and 50° W. Eos, 84:OS80
Bisagni, J. J., H-S. Kim, and K. F. Drinkwater (submitted) Observations and modeling of shelf-slope front seasonal
variability between 75° and 50° W. Deep-Sea
Research II.
JJB-4: Research Experiences for
Undergraduates (REU) Supplement to Award No. ANI-0124945 (Internet2 Connection for the
Avijit
Gangopadhyay: Award No. ANI-0124945 (Internet2 Connection for the University of
Massachusetts Dartmouth’s School for Marine Science and Technology,
(Co-investigator;
W. Brown, Principal Investigator), NSF, 6/1/2002-9/30/2003, $150,000. provided
expertise to help set up Internet2 technology at SMAST for large datasets and
model output transfer between modeling groups at SMAST, Harvard, JPL/NASA and
JHU/APL.
Dian J. Gifford: (DJG)-1: Collaborative Research: GLOBEC-01:Patterns of energy flow
and utilization on Georges Bank, Principal
Investigator: D. Gifford, with J. Steele, J. Bisagni, J. Collie, E. Durbin, B.
Sullivan, M. Sieracki, M. Fogarty, D. Mountain, J. Link and D. Palka, Co-Is.
OCE 0217399. 2002-2006, $507,382
(URI component). Results summarized
in Bisagni’s section above.
DJG-2: Collaborative Research: Initiation and Maintenance of Population Maxima
of the Ctenophore Mnemiopsis leidyi in Northern Coastal Waters.
Co-Investigator with B.K. Sullivan, Principal Investigator and J.H. Costello,
Co-Investigator. OCE 0115177 (BKS and DJG)
($655,401), 2001-2004. In situ rate processes relevant to ctenophore population
dynamics and bloom initiation and maintenance were monitored weekly at three
stations for two complete seasonal cycles, together with actual population
response to changes in ambient physical and biological parameters. New and
compelling hypotheses were developed concerning conditions that are critical
for triggering rapid population growth of M.
leidyi. Hydrographic regimes
that determine population distributions of M.
leidyi during the winter months prior to bloom initiation, transport to
regions where ctenophore blooms are initiated in spring, and periods of intense
warming and water column stratification all appear to control the timing and
magnitude of ctenophore blooms. Significant revision of our previous understanding
of the species’ biology includes new insight into physiological limits,
particularly feeding and reproduction at low temperature. Plankton distribution
and abundance data have been added to the URI Plankton Time Series web site
(http://www.gso.uri.edu/phytoplankton/). Three graduate and 6 undergraduate
students were supported by the award. Presentations were made at the ASLO
summer meeting, Victoria, BC (2002), ICES 3RD International
Zooplankton Production Symposium, Dijon, Spain (2003) (3 presentations), and
ASLO winter meeting, Salt Lake City, UT (2003). Publications to date resulting
from the research:
Sullivan, Lindsay J, and D.J. Gifford (2004) Diet of the larval
ctenophore Mnemiopsis leidyi
A.Agassiz (Ctenophora, Lobata). J. Plankton Res. 26: 417-431.
Costello, J.H. and H.W. Mianzan. 2003. Sampling field distributions of Mnemiopsis leidyi (Ctenophora, Lobata):
planktonic or benthic methods? J. Plankton Res. 4: 455-459.
Costello, J.H., B.K. Sullivan, D.J. Gifford, D. VanKeuren and L.J. Sullivan.
Seasonal refugia, shoreward thermal amplification and metapopulation dynamics
of the ctenophore Mnemiopsis leidyi in
Sullivan, B.K., J.H, Costello and D.J. Gifford. A mechanism linking climate change and
trophic mismatch in coastal plankton. Nature.
Submitted.
1. Introduction, Motivation, and
Background
This proposal addresses several mechanisms by which remote
(basin-scale) forcing is likely impact the population dynamics and production
of Calanus finmarchicus in the coupled Georges Bank/Gulf of
In order to
address these objectives, we will:
(i)
analyze selected satellite and in-situ
data sets collected during the GLOBEC Georges Bank period;
(ii)
simulate basin-scale circulation fields for the 1990s GLOBEC study
period using an eddy-resolving ROMS already in use and under validation, and
(iii)
conduct a series of coupled biophysical numerical experiments to
test a set of hypotheses on the remote forcing of C. finmarchicus supply and productivity in the NWA/GB system.
We will use newly compiled datasets on nutrient
concentrations and Calanus abundances from the western
1.1 Life History of Calanus finmarchicus. The early life history stages and copepodids of mesozooplankton,
especially copepods like Calanus
finmarchicus, Pseudocalanus spp.
and Centropages sp., are major prey for the larvae of
economically valuable, historically harvested groundfish species (cod, haddock)
on Georges Bank. In early spring, C. finmarchicus are by far the dominant
component of mesozooplankton biomass on
1.2 A Spawning Stock Size Approach to Calanus finmarchicus. For a
century, fisheries biologists have documented strong impacts of fish stock
sizes on subsequent recruitment (the so-called stock-recruit relationship). We believe such an approach is equally
valuable in considering interannual variability in C. finmarchicus
population dynamics and abundances.
A general view of the
1.3 Impact of Climate-related Variability on Calanus finmarchicus. In
the Northeast Atlantic there is substantial evidence that Calanus finmarchicus population abundances have varied inversely
with the North Atlantic Oscillation (Planque and Reid, 1998; Greene et al. 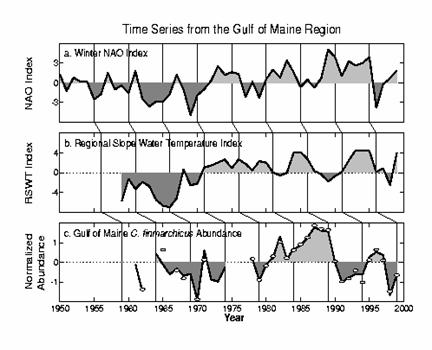 2003), but C. finmarchicus abundance in the Gulf of
Maine during 1960-90 covaried directly with the NAO (Conversi et al. 2001), suggesting that perhaps
different mechanisms, acting through temperature and ocean circulation
patterns/intensities are operating on the eastern and western sides of the
North Atlantic. However, Planque
and Reid (1998) also document the sometimes ephemeral nature of such
climate-ecosystem relationships, as they observed the pattern for the
2003), but C. finmarchicus abundance in the Gulf of
Maine during 1960-90 covaried directly with the NAO (Conversi et al. 2001), suggesting that perhaps
different mechanisms, acting through temperature and ocean circulation
patterns/intensities are operating on the eastern and western sides of the
North Atlantic. However, Planque
and Reid (1998) also document the sometimes ephemeral nature of such
climate-ecosystem relationships, as they observed the pattern for the  salinity variability on the
Scotian Shelf and in the
salinity variability on the
Scotian Shelf and in the
There
was no apparent immediate and dramatic response of the plankton community in
the GoM/GB region to the 1995-96 NAO reversal. Bisagni et al. (2001) document significant cooling of adjacent slope water
surface temperatures (using AVHRR) from 1994-1996, which covaried with volume
transport anomalies through the Northeast Channel. Calanus finmarchicus populations on Georges Bank had slightly
slower development and later maturation times due to lower temperatures in 1996
than in 1995 (Durbin et al.
2000).
More
significant impacts on the plankton community were observed in the summer of
1998. During
the 1970s, ‘80s, and early ‘90s, conditions over the Scotian Slope
and in the deep basins of the Scotian Shelf and
1.4 Mechanisms for Climate-related Variability of Calanus Spawning Stock
Size in NWA. Interdecadal variability (e.g., 1960s cold
anomaly; warm 1970s) in Western Boundary Current (WBC) transports (Petrie and
Drinkwater 1993; Drinkwater et al.
unpubl.) can create large changes in ocean temperature, particularly in deeper
waters, that alter the distribution, migrations, spawning times, growth and
physiological rates of resident organisms.
The massive kill of tilefish along the Mid-Atlantic Bight in the early
1880s may be an extreme example of the impacts from an excessive inflow of cold
subpolar water (Marsh et al. 1999).
Altered temperature is likely to impact the physiology of diapausing C. finmarchicus and disrupt the timing
of diapause exit, for which the cues are unknown. If so, this could impact the synchrony
of C. finmarchicus return to the
surface relative to the seasonal production cycle and phytoplankton blooming.
Increased equatorward advection along the shelf-slope should provide enhanced
southward transport of organisms, and this may lead to export of Calanus finmarchicus from the Labrador
Sea to the
1.5
Multi-Decadal to Interannual Variability of Basin-scale Circulation Related to
NAO. Recent studies (Hoerling et al. 2001; Hurell et al., 2001) show that the NAO has a major impact on the North Atlantic circulation and variability. Taylor and
Stephens (1998) show that over the period 1966-96, the position of the Gulf
Stream north wall (GSNW) appears to be determined by the NAO, but with a lag of
about 2 years. They attribute the lag to propagating Rossby waves (Gangopadhyay
et al. 1992), and suggest that the
delayed-response predictability observed for the 1977 to 1988 period has been a
feature of the Gulf Stream system for last 30
years. Figure 2 shows the Gulf Stream position
(position of the 15oC isotherm from data in Parker et al., 1995) overlaid with winter NAO
from Drinkwater (pers. comm.). Both Gulf Stream
position and NAO index exhibit an upward trend since 1970 (Conversi et al. 2001; Taylor and Gangopadhyay
2001). There was a downward trend from 1950 to 1970, suggesting that these
trends may be due to climate variability. The Gulf Stream
was furthest south during the 1960s, and furthest north during the 1950s and
1980s. The Gulf Stream position exhibits
pronounced interannual variation at periods of 3-5 years.
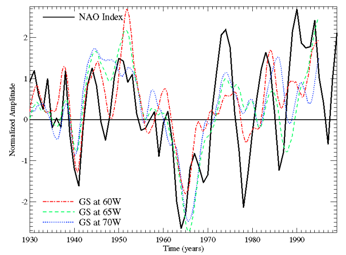
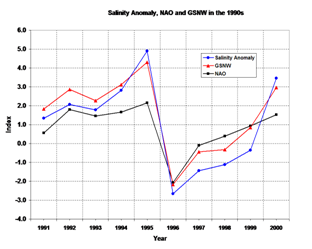

 Fluctuations of the
Fluctuations of the
Fratantoni and Pickart (2005, submitted ms.) analyzed twelve years
(1990-2001) of hydrographic data to examine alongstream evolution of the
shelfbreak front in the western north Atlantic from the west coast of Greenland
to the Mid-Atlantic Bight.. They found that the shelfbreak front in the
Labrador Sea is composed of two fronts: one inshore front with fresher Labrador
Sea water, and the other offshore with
These three studies raise questions regarding the source of the
low-salinity waters in the
1.6 Ongoing Basin-scale Modeling for Simulating High and Low NAO Forced
Fields.  The physical modeling
component is based on ROMS (Regional Ocean Modeling System), which was
developed and modified by
The physical modeling
component is based on ROMS (Regional Ocean Modeling System), which was
developed and modified by
Using NASA Interdisciplinary Science
(IDS) funding (NASA NNG04GH50G), we purchased an Altix 350, eight-processor SGI
computer for ROMS modeling. The eddy-resolving 15-20km ROMS has been configured
for the  For NASA, we are carrying out two
10-year-long simulations representative of the two phases of the NAO. We have
started the simulation for the high-NAO period. The initial
temperature-salinity fields are from Levitus and the forcing fields are based
on the annual climatology, computed for 1980-93, by the Southampton
Oceanographic Center (SOC). The model is (as of
For NASA, we are carrying out two
10-year-long simulations representative of the two phases of the NAO. We have
started the simulation for the high-NAO period. The initial
temperature-salinity fields are from Levitus and the forcing fields are based
on the annual climatology, computed for 1980-93, by the Southampton
Oceanographic Center (SOC). The model is (as of
In summary,
studies have shown correlations among the basin-scale NAO index and the
circulation variability realized by Gulf Stream position,
2. Research Objectives,
Questions & Hypotheses
2.1 Climate-Related Basin-Scale Circulation
Variability Questions, Two distinct time-scales are important
for evaluating the impact of climate and basin-scale forcing on the GoM/GB
ecosystem. The first is the multi-decadal variability (Petrie and Drinkwater
1993) which results from the accumulated effects of a sustained high-NAO or
low-NAO condition. Clearly, the C. finmarchicus population responds to
sustained long-term changes in the system. Our present NASA modeling effort will address this with
the addition of biological modeling.
Rossby (1999) attributes the annual north-south
shifting of the eastward-flowing
We will peform data analysis, modeling, and diagnostic
analysis, to address several scientific questions about the NAO and its impact
on western
(1) What is the
interannual variability of the kinematical relationship between features such
as the north wall of the Gulf Stream,

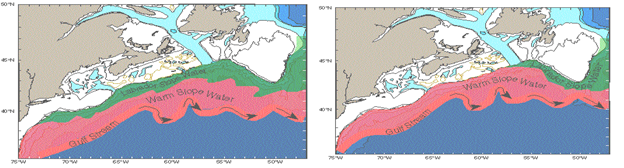 (2) What is the
nature of the variability of these features during the 1993-1999 US-GLOBEC
measurement period? It is presently unclear how the salinity anomaly occurred
during the 1990s. The high correlation between the freshening of the GoM/GB
region with southward movement of the
(2) What is the
nature of the variability of these features during the 1993-1999 US-GLOBEC
measurement period? It is presently unclear how the salinity anomaly occurred
during the 1990s. The high correlation between the freshening of the GoM/GB
region with southward movement of the
(3) What is the nature of the
interactions between the subtropical gyre and the subpolar gyre in simulations
conducted for high-NAO years versus low-NAO years? Their interaction through
the deeper waters in the thermohaline circulation may be very important for
determining the fate of Calanus finmarchicus being
transported from
2.2 Climate-Related C.
finmarchicus Variability Questions. A goal of the proposed
research is to examine the impacts of seasonal and interannual variation in
ocean conditions in the western North Atlantic (Labrador Sea, 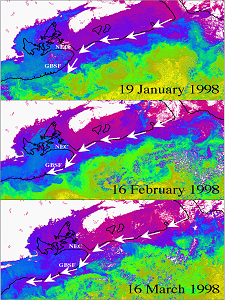 (Figure 1). This is counter to expectations; given
the high C. finmarchicus abundances
in both surface layers (during spring-summer) and deep diapausing layers
(remainder of year) in the Labrador Sea, one might expect higher C. finmarchicus seed supply when
Labrador influences are great in the Slope Sea region. The discrepancy may be
related to temperature of the water that the diapausing stocks are in, and the
role that temperature-controlled physiological processes have on terminating the
diapause stage. Head et al. (2000)
report that surface waters of the
(Figure 1). This is counter to expectations; given
the high C. finmarchicus abundances
in both surface layers (during spring-summer) and deep diapausing layers
(remainder of year) in the Labrador Sea, one might expect higher C. finmarchicus seed supply when
Labrador influences are great in the Slope Sea region. The discrepancy may be
related to temperature of the water that the diapausing stocks are in, and the
role that temperature-controlled physiological processes have on terminating the
diapause stage. Head et al. (2000)
report that surface waters of the  Atlantic. We will test the hypothesis that
basin-scale climate oscillations (NAO), operating through time-varying
intensity of deep and intermediate water-mass formation and ocean circulation,
result in shelf- and slope-water transports and fluxes of Calanus and nutrients that are advantageous at some times and
disadvantageous at others for the development of large overwintering
populations of C. finmarchicus in
deep waters adjacent to the GB/GoMsystem.
Atlantic. We will test the hypothesis that
basin-scale climate oscillations (NAO), operating through time-varying
intensity of deep and intermediate water-mass formation and ocean circulation,
result in shelf- and slope-water transports and fluxes of Calanus and nutrients that are advantageous at some times and
disadvantageous at others for the development of large overwintering
populations of C. finmarchicus in
deep waters adjacent to the GB/GoMsystem.
The
core hypothesis and predictions of this work are:
Hypothesis: The
occurrence of large populations of Calanus
finmarchicus in the coupled GB/GoM system REQUIRES (1) high seed stocks
(supply) of diapausing C.finmarchicus
in the deeper ocean regions nearby (GOM basins and the Slope Sea), (2) that the
deep C. finmarchicus stocks terminate
diapause at the appropriate time to be synchronous with continental shelf
spring blooms, and (3) a nutrient enriched, highly productive ecosystem in the GB/GoM
to sustain high growth and survival rates of Calanus that will provide seed for the subsequent year.
Prediction
A: Overwintering Calanus
finmarchicus seed stocks are LOW and GB/GoM productivity is HIGH when the
water masses of the Slope Sea have little influence (input) from
Labrador-Irminger Gyre (Labrador Slope Water) water masses (due to the
relatively nutrient replete bottom waters and low Calanus supply in Warm Slope Waters), but C. finmarchicus recruitment is good because of a near-perfect match
between the time of diapause awakening and the time of the spring bloom, the
latter of which is large because of the higher concentration of nutrients in
deep warm slope waters.
Prediction
B: Overwintering C. finmarchicus seed stocks are HIGH
and GB/GoM productivity is LOW when the water masses of the Slope Sea have a
large proportion of Labrador Sea water (due to the relatively nutrient-depleted
bottom waters and high C. finmarchicus
supply in cold Labrador Slope Water), but
recruitment and productivity are poor because of the generally low
springtime productivity (low nutrients) and a timing mismatch between diapause
awakening, ascent and reproduction and the NW Atlantic spring bloom.
3. Research Approach
It is clear that the relationship between the GB/GoM ecosystems
and basin-scale variability related to NAO occurs over at least two different
time-scales: (i) multi-decadal, and (ii) interannual. We propose to carry out a
set of eddy-resolving basin-scale model simulations, analyze satellite and
other available data sets from the GLOBEC period, validate the model
simulations, and use the simulations to address a set of questions that relate
ecosystem variability in the Scotian Shelf,
Our approach is based on (i) analyzing satellite and
hydrographic observations during 1993-1999 period; (ii) simulating the
basin-scale impacts on Calanus finmarchicus
from an eddy-resolving North Atlantic model simulation of high vs. low NAO
periods; (iii) simulating basin-scale physical and biological fields for the
GLOBEC period (1993-1999); and (iv) synthesizing the model simulations and
GLOBEC data sets during the 1993-1999 period to address the specific questions
outlined in Section 2 above. In addition, we will provide the basin-scale
simulation fields for forcing other regional models (both physical and
biophysical) to specifically address detailed hypothesis related to the
interannual variability of the GB/GoM ecosystems.
3.1 Analysis and Use of
Satellite-derived SST (1993-1999). We propose a
comprehensive synthesis using both daily satellite-derived SST data from the
western
3.2 Basin-Scale Circulation
Modeling
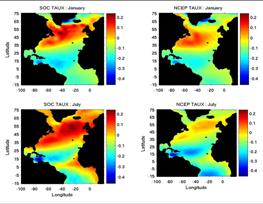 . Two of us (AG and JB) are funded by NASA to simulate
decade long periods of high and low NAO conditions. The high NAO conditions are currently
being simulated. During summer
2005, simulation for the Low-NAO period will be done. Levitus will be used for
initial temperature and salinity fields. Wind forcing is derived from the
ICOADS data set during the sixties (1962-1971, low-NAO period). Zonal
wind-stress fields for winter (January) and summer (July) for high and low-NAO
periods are shown in Figure 7. We
are addressing a hypothesis related to ‘depletion vs. dilution’ of
nutrients, i.e., the dominance of low nutrient Labrador Sea Water influx in the
Slope Sea during low-NAO periods as opposed to high nutrient warm slope water
coming through the Gulf Stream system during high-NAO periods. So, we focus the results of the
simulations towards realistically generating the Gulf Stream System and the
Labrador Sea Water influx into the
. Two of us (AG and JB) are funded by NASA to simulate
decade long periods of high and low NAO conditions. The high NAO conditions are currently
being simulated. During summer
2005, simulation for the Low-NAO period will be done. Levitus will be used for
initial temperature and salinity fields. Wind forcing is derived from the
ICOADS data set during the sixties (1962-1971, low-NAO period). Zonal
wind-stress fields for winter (January) and summer (July) for high and low-NAO
periods are shown in Figure 7. We
are addressing a hypothesis related to ‘depletion vs. dilution’ of
nutrients, i.e., the dominance of low nutrient Labrador Sea Water influx in the
Slope Sea during low-NAO periods as opposed to high nutrient warm slope water
coming through the Gulf Stream system during high-NAO periods. So, we focus the results of the
simulations towards realistically generating the Gulf Stream System and the
Labrador Sea Water influx into the
3.2.1 Proposed basin-scale simulations. In the first year, we will integrate the North Atlantic ROMS OGCM
from its final high-NAO state with monthly NCEP reanalysis fluxes during
1988-1999. Based on our experience (Chao et al., 1996), we start the simulation
five years before the start of GLOBEC (1993) is to allow the winds to force the
response of the


 The success
criteria for the proposed physical and biochemical simulation will be our
ability to: (i) reproduce the high-NAO vs. low-NAO behavior of the GS path
similar to that shown in Fig. 4, (ii) reproduce the nutrient variability in the
The success
criteria for the proposed physical and biochemical simulation will be our
ability to: (i) reproduce the high-NAO vs. low-NAO behavior of the GS path
similar to that shown in Fig. 4, (ii) reproduce the nutrient variability in the
We will investigate transport pathways from the Labrador Sea/Slope
to the
3.2.2 High-resolution field generation using
basin-scale output and GLOBEC data sets. We propose to generate a series of
high-resolution (5-km) fields by melding the basin-scale fields (10-15 km
resolution) with GLOBEC period data sets using multi-scale objective analysis
(OA) as done previously ((Gangopadhyay et
al. 2002, 2003). The OA is performed in two stages (Lermusiaux, 1999a). In
the first stage, the largest dynamical scales (from the basin-scale model
fields) are resolved at each level, using estimated large-scale e-folding
spatial decays, zero crossings, and temporal decay. In the second stage, the
synoptic dynamics of interest (meso-scale and sub-basin scale, from data or
synthetic profiles as explained below) are resolved using its estimated
space-time decays. The background for this second stage OA is the first-stage
OA. The primary assumption made in this two-scale OA is that the errors in the
basin-scale (first-stage) and synoptic (second-stage) dynamical scales are
statistically independent. This procedure effectively and smoothly melds the
synoptic profiles or observational profiles and climatology for the GoM/GB
region. For detail mathematical description of the OA parameters, see
Lermusiaux (1999a,b).
The high-resolution synthesis will be carried out in four
steps. First, the temperature and salinity fields from the basin-scale ROMS
simulation will be averaged over 5-day periods during the GLOBEC period of
1993-1999. The locations for these profiles are shown in Figure 9 as black dots
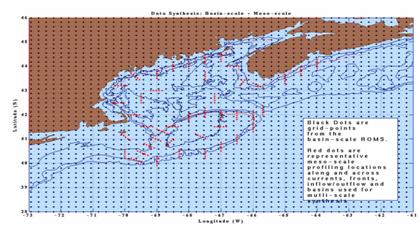 within
the region of interest (73-50W, 38-46N). Second, the 5-day averaged SST fields
and the ROMS temperature field at the bottom level will be synthesized to
develop synthetic temperature profiles at pre-determined locations (red dots in
Figure 9). The synthesis equation is given by: T(z) = Ts - (Ts
– Tb) Ф(z). Here Ts is the surface
temperature (SST) and Tb is the bottom temperature from
within
the region of interest (73-50W, 38-46N). Second, the 5-day averaged SST fields
and the ROMS temperature field at the bottom level will be synthesized to
develop synthetic temperature profiles at pre-determined locations (red dots in
Figure 9). The synthesis equation is given by: T(z) = Ts - (Ts
– Tb) Ф(z). Here Ts is the surface
temperature (SST) and Tb is the bottom temperature from  the nearest ROMS
grid point. Ф(z) is the non-dimensional structure function (unity at surface to
zero at bottom) for the temperature profile of the mesoscale feature at the
location of interest. The forms of Ф(z) for all the 270 profiles are
available from Gangopadhyay et al.
(2003). Third, the corresponding salinity profiles will be determined using
appropriate GLOBEC data-derived T-S relationships in conjunction with the T(Z)
derived in step 2. Finally, the mesoscale temperature and salinity profiles
from above two steps will be melded with the basin-scale fields for each five
day period using multiscale OA. This process will generate three-dimensional
high-resolution fields for temperature, salinity and geostrophic velocity every
five-days during the whole GLOBEC period (1993-1999) that has the climatic and
basin-scale impact of NAO in the flow field. We will thus resolve the
sub-mesoscale variability of the Scotian Shelf and the flow through the
Northeast channel into the
the nearest ROMS
grid point. Ф(z) is the non-dimensional structure function (unity at surface to
zero at bottom) for the temperature profile of the mesoscale feature at the
location of interest. The forms of Ф(z) for all the 270 profiles are
available from Gangopadhyay et al.
(2003). Third, the corresponding salinity profiles will be determined using
appropriate GLOBEC data-derived T-S relationships in conjunction with the T(Z)
derived in step 2. Finally, the mesoscale temperature and salinity profiles
from above two steps will be melded with the basin-scale fields for each five
day period using multiscale OA. This process will generate three-dimensional
high-resolution fields for temperature, salinity and geostrophic velocity every
five-days during the whole GLOBEC period (1993-1999) that has the climatic and
basin-scale impact of NAO in the flow field. We will thus resolve the
sub-mesoscale variability of the Scotian Shelf and the flow through the
Northeast channel into the
3.2.3 Multi-decadal and Interannual Validation of Simulations. The model simulations will be
validated with data sets available for the
3.3 The C. finmarchicus Modeling
Experiments. The biological modeling follows naturally from the analysis of the
physical fields and will be built incrementally from that foundation. The population dynamics of Calanus finmarchicus and transports by
oceanic flows have been extensively modeled in both the Northwest Atlantic
(Miller et al. 1998) and other
regions of the North Atlantic (Tittensor et al. 2003; Bryant et al. 1998; Carlotti and Wolf, 1998; Carlotti and Radach 1996;
Harms et al. 2000). However, almost all of those prior
efforts have emphasized the population dynamics of the C. finmarchicus when they are actively feeding and reproducing in
the near-surface layers. An
emphasis of the coupled biophysical Lagrangian modeling we propose is to
examine the role of transports on supply of deep diapausing populations of C. finmarchicus to the Slope Sea region
from the Labrador Sea, where diapausing populations are very large (>17000 m-2)
at depths of 300-800m (Heath et al.
2004; Head, pers. comm.). The
Labrador Sea is recognized as one of the epicenters of C. finmarchicus in the North Atlantic (Tittensor et al. 2003; Heath et al. 2004), so export of water from the Labrador
Sea to regions further south would be expected to provide influx
of Calanus. Transports of C. finmarchicus from diapausing populations to the North Sea have
been the focus of both field studies and extensively modeled (Heath 1999; Heath
et al. 1999; Heath and Jonasdottir
1999; Gallego et al. 1999), but our
proposed modeling is the first to examine transport of diapausing populations
in the western North Atlantic .

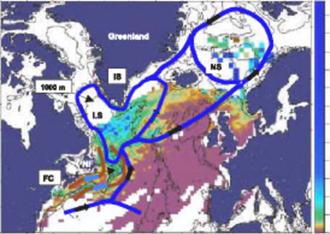 There are three
distinct biological (Calanus finmarchicus)
model experiments, which are linked directly to the physical model
experiments. The first experiment
(done in year 1) will use the results of the decade long simulations of the
“prototypical” high NAO (1980-93) and low NAO (1962-71)
periods. Basin-scale physical
simulations for both high and low NAO periods are currently being done (with
NASA funding), so the velocity and tracer fields will be available in year one
for Lagrangian modeling of Calanus
transports and physiology. The
second biological modeling experiment (year 2), will use velocity and tracer
fields from simulations of the GLOBEC field years (1993-1999) to explore the
impact of the 1996 NAO reversal and later inflow of Labrador Slope Water into
the NW Atlantic on diapausing Calanus
finmarchicus populations.
Comparison of the results of multiyear model simulations from experiment
1 and experiment 2 will provide insights on the effects of a prolonged influence of cold, nutrient
deplete Labrador water on the
There are three
distinct biological (Calanus finmarchicus)
model experiments, which are linked directly to the physical model
experiments. The first experiment
(done in year 1) will use the results of the decade long simulations of the
“prototypical” high NAO (1980-93) and low NAO (1962-71)
periods. Basin-scale physical
simulations for both high and low NAO periods are currently being done (with
NASA funding), so the velocity and tracer fields will be available in year one
for Lagrangian modeling of Calanus
transports and physiology. The
second biological modeling experiment (year 2), will use velocity and tracer
fields from simulations of the GLOBEC field years (1993-1999) to explore the
impact of the 1996 NAO reversal and later inflow of Labrador Slope Water into
the NW Atlantic on diapausing Calanus
finmarchicus populations.
Comparison of the results of multiyear model simulations from experiment
1 and experiment 2 will provide insights on the effects of a prolonged influence of cold, nutrient
deplete Labrador water on the
For
each circulation scenario, we will begin with an analysis of the transport
using tracer experiments with Lagrangian models. Note that the high vertical
resolution (50 levels) of the present ROMS set up will provide more accurate
vertical fluxes than many studies before. Biological realism will be added
sequentially. Deep diapausing C.
finmarchicus populations in the southern Labrador Sea will be initiated
with a range of lipid stores; timing of awakening will be related to lipid
consumption at depth (recognizing that most lipid is not consumed during the
diapause phase, but rather remains available for egg production immediately
following maturation) (Irigoien, 2004), and therefore a function of
temperature. Heath et al. (2004) inferred from the assumed
neutral buoyancy depth of diapausing C5 C. finmarchicus in the
The C. finmarchicus
biological simulations (experiments 1-3) are complementary to already funded Calanus modeling projects (Franks et al. project) that are more specific
to the dynamics of Calanus during the
active season in the GoM/GB and Scotian Shelf regions. Our project focuses on the transports
and physiological awakening of Calanus
during their deep-dwelling diapause phases and on the implications of NAO
variability and cold
Significance and Intellectual
Merit. This project will develop a quantitative understanding
of how
To address variability over these multiple time scales,
we propose to do the following during the two-year duration of the project: (1)
Set up and run an individual based model (IBM) for the Northwest Atlantic,
using the high-NAO (1980-1993) and low-NAO (1962-1971) forced physical fields
from an ongoing eddy-resolving North Atlantic simulation to understand
multi-decadal variability of Calanus
finmarchicus seeding and production in this region; (2) Perform a set of
eddy-resolving basin-scale model simulations during 1988-1999 starting from
already existing high-NAO simulations (from an ongoing NASA project) and run
the IBM to study the interannual variability of C. finmarchicus seeding and production in this region; (3) Analyze
long-term in-situ physical and
biological datasets and satellite-derived sea surface temperature (SST) along with
in-situ physical, biological, and
chemical data collected during the GLOBEC core-measurement period (1995-1999),
and validate the two basin-scale physical and biological fields to develop a
broader understanding of C. finmarchicus
seeding and production; and (4) Generate four-dimensional high-resolution
(5-km) physical fields using basin-scale fields and available data during
1993-1999, and run a series of IBM simulations at higher resolution to address
questions relating ecosystem variability on the Scotian Shelf, on Slope Sea and
within the Gulf of Maine and on Georges Bank to the large-scale fluctuations of
the NAO.
Broader Impacts. We
anticipate that our work with satellite-derived SST data under this proposal
will provide a synthetic focus to assist with other regional observational and
modeling work being proposed by other groups. Our
study on the impact of large-scale climatic and basin-scale forcing on the
regional ecosystem of the GoM/GB region will enhance the understanding of the
larger community. We will train two graduate students during the two years of
this project. Results will be disseminated by peer-reviewed scientific
publications and presentations at national conferences. We will make the model
output available via a website linked to the GLOBEC website. We will give
public lectures at