PROJECT DESCRIPTION
I. Results from Prior NSF Support
Title:
Zooplankton
and Nekton in the
Principal Investigators: Peter Wiebe, Timothy Stanton, and Charles Greene
Project Duration: 3/1/95-2/28/97 Award Amount: $399,477
Title:
populations from the
Principal Investigators: Charles Greene, Mark Benfield, and Peter Wiebe
Project Duration: 1/1/97-6/30/01 Award Amount: $645,846
Title:
in Relation to Micro-, and Coarse-scale Physical Structure in the Georges Bank Region (supplement to NSF OCE-9313675)
Principal Investigators: Peter Wiebe, Timothy Stanton, and Charles Greene
Project Duration: 2/15/98-1/31/99 Award Amount: $90,000
In addition
to the above grants, Charles Greene was supported during 2000/2001 as a
sabbatical fellow at the
The most
significant scientific result derived from the above research has been our
ability to link the order of magnitude reduction in C. finmarchicus abundance observed in the Gulf of Maine during
autumn 1998 (relative to autumns of 1997 and 1999) to a North Atlantic
Oscillation (NAO)-driven modal shift in the Northwest Atlantic’s coupled
slope water system (Greene and Pershing, 2000; MERCINA, 2001; 2003; 2004). Retrospective analyses of continuous
plankton recorder and hydrographic time-series data have enabled us to place
this result in the context of climate-driven changes in ocean circulation observed
over the past 50 years in the
A. Special Symposia and Workshops:
The
Response of Northeast and
Response
of
Marine
Ecosystem Responses to Climate: The Responses of Large Marine Ecosystems to
Interdecadal-Scale Climate Variability. ASLO/AGU Ocean Sciences Meeting,
B. Invited Talks at International Symposia:
Greene,
C.H., and A.J. Pershing. The
response of Calanus finmarchicus
populations to climate variability in the
Greene,
C.H., and A.J. Pershing.
Trans-Atlantic responses of Calanus
finmarchicus to basin-scale forcing associated with the North Atlantic
Oscillation. AGU Chapman Conference on the North Atlantic Oscillation,
Greene,
C.H., and A.J. Pershing.
Trans-Atlantic responses of Calanus
finmarchicus to basin-scale forcing associated with the North Atlantic
Oscillation. 70th Anniversary of the Continuous Plankton Recorder
Surveys of North Atlantic Symposium,
Greene,
C.H., and A.J. Pershing. Biocomplexity and climate:
recovery of the North Atlantic Right Whale population in the context of
climate-induced changes in oceanographic processes. Climate Change and
Aquatic Systems Symposium,
C. Publications Citing Previous US GLOBEC
Support:
The
following 20 publications cite previous US GLOBEC support through the above
grants: Barton et al. (2003); Benfield et al. (1998, 2003); Drinkwater et al.
(2002); Greene et al. (1998a, b, c, d, 2003); Greene
and Pershing (2000, 2003, 2004); MERCINA (2001, 2003, 2004); Pershing et al.
(2004, 2005); Wiebe et al. (1996, 1999, 2002).
D. Educational Outcomes from Previous
US GLOBEC
provided support for the following graduate thesis research: Andrew Barton
(Cornell MS 2001: Continuous plankton
recorder survey phytoplankton measurements and the North Atlantic Oscillation:
interannual to multidecadal variability in the Northwest Shelf, Northeast
Shelf, and Central North Atlantic Ocean), Andrew Pershing (Cornell PhD
2001: Response of large marine ecosystems
to climate variability: patterns, processes, concepts, and methods), Karen
Fisher (Cornell PhD 2002: Intermittency
of spatial and temporal plankton patterns), and Joseph Warren (WHOI/MIT PhD
2001: Estimating Gulf of Maine
zooplankton distributions using multiple frequency acoustic, video and
environmental data). Seven Cornell undergraduates conducted related
research in our laboratory during this time frame, resulting in two honors
theses.
II. Proposed Research
A. Introduction
Understanding the variability
observed in marine ecosystems, especially the large fluctuations in abundance
and recruitment of exploited fish stocks, has been a major goal of oceanographers
and fisheries scientists since the late 19th century (reviewed by Cushing,
1982; 1996). For many years,
researchers have noted that fluctuations in fish populations are often
associated with dramatic regime shifts in marine ecosystems (de Young et al.,
2004). One of the earliest
recognized and most famous of these regime shifts is the Russell Cycle, which was first observed as a shift in the English
Channel's plankton community during the late 1920's that was subsequently
observed to reverse itself during the late 1960's (Cushing and Dickson, 1976;
Cushing, 1982). The Russell Cycle was characterized by
dramatic changes in nutrient concentrations as well as the species composition
and relative abundances of phytoplankton, zooplankton, and fish. A key commonality between the Russell Cycle and other fluctuations
observed in North Atlantic fisheries is their link to basin-scale changes in
The major
basin-scale mode of inter-annual to inter-decadal climate variability in the
In addition to the NAO, there
are other natural and anthropogenic sources of basin-scale climate variability
in the
B. Hypotheses
Until very
recently, many oceanographers and climate scientists have expressed serious
concerns about the potential for abrupt climate change and dramatic cooling in
northern
1.
Remote forcing of ecosystem processes in the
SS/GOM/GB region is mediated not only by the CSWS but also through the enhanced
transport of lower salinity shelf waters derived from upstream sources,
including the
2.
Remote forcing from the
C. Research Plan/Methods
In describing our research plan, we will detail what we have learned from previous synthesis studies about regional responses to climate variability from the SS to GB and the methods that we used to achieve this knowledge. Since our hypotheses arose in the context of these previous studies, this comparative approach should provide the reviewer with a clearer understanding of what we hope to learn in the coming years and the methods that we propose to employ.
Hypothesis 1: Remote forcing of
ecosystem processes in the SS/GOM/GB region is mediated not only by the CSWS
but also through the enhanced transport of lower salinity shelf waters derived
from upstream sources, including the
The region
of the Northwest Atlantic shelf extending from the SS to GB lies within a
shifting oceanographic transition zone, located between cold subpolar waters
influenced by fluctuations in the Labrador Current to the northeast and warm
temperate waters influenced by fluctuations in the Gulf Stream to the south
(Loder et al., 2001; MERCINA, 2001). The transitions that occur within this
zone are not only physical, as reflected by hydrographic changes, but also
biological, as reflected by changes in the composition and relative abundance
of plankton (Greene and Pershing, 2000; MERCINA, 2001; 2003; Sameoto, 2001).
The shifting nature of this transition zone makes the region especially
vulnerable to climate-driven changes in
The
region’s physical responses to changes in ocean circulation are often
mediated by the
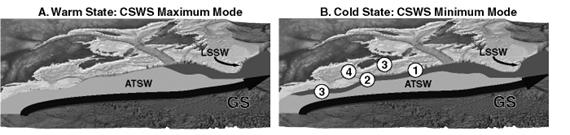
Figure 1.
Distribution of LSSW (dark gray) and ATSW (light gray) during the maximum (A)
and minimum (B) modal states of the CSWS.
The circled numbers indicate the observations of LSSW following the 1996
NAO event: 1 = September 1997, 2 =
January 1998, 3 = February 1998, 4 = August 1998. Redrawn from Greene and
Pershing (2003).
Recently, it has been shown that modal shifts in the CSWS are often associated with phase changes in the NAO (MERCINA, 2001). From 1975 to 1999, the NAO Index was predominantly positive (Hurrell et al., 2003), and the CSWS usually exhibited conditions characteristic of its maximum modal state (Fig. 2A, B). However, five times during these 25 years (1977, 1979, 1985, 1987, 1996), the NAO Index dropped to negative values for a single year. In each case, the CSWS responded to a drop in the index by shifting toward its minimum modal state after a one- to two-year time lag (1978, 1981, 1987, 1989, 1998). While the first two responses of the CSWS, in 1978 and 1981, were relatively small, the latter responses were more substantial. The response to the 1985 and 1987 drops in the NAO Index involved a multi-year modal shift lasting from 1987 to 1990. The response to the 1996 drop in the index was the largest and best documented modal shift to date.
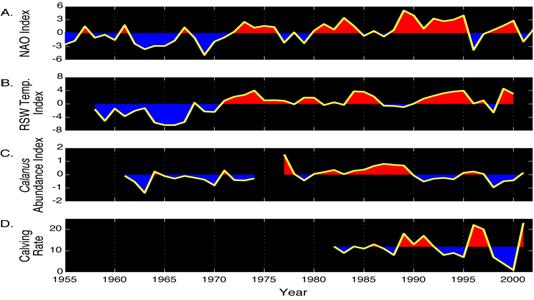
Figure 2. Time series from the
B.) Annual
values of the Regional Slope Water (RSW) Temperature Index. C.) Annual values of the Calanus
finmarchicus Abundance Index. D.
Annual values of right whale calving rate. The winter NAO Index is the mean
atmospheric pressure difference between the North Atlantic’s subtropical
high-pressure system, measured in
In 1996, the NAO Index exhibited its largest single-year drop of the 20th century, attaining a negative value not seen since the 1960’s (Fig. 2A). This large drop in the NAO Index was followed over the next two years by a modal shift in the CSWS, with the Labrador Current intensifying and the LSSW steadily advancing along the shelf break, displacing ATSW offshore, and penetrating farther southwest into the MAB (Fig. 1A) (MERCINA, 2001). In addition to its advance along the shelf break, the LSSW also invaded the shelf waters of the SS, GOM, and GB. Cooler temperatures and lower salinities were observed throughout the region in 1998, especially in the deep-basin waters derived directly from slope-water incursions.
The hydrographic changes observed in the region during 1998 were short-lived, however. The large drop in the NAO Index during the winter of 1996 was a single-year event, and the Index returned to positive values for the remainder of the 1990’s. Similarly, the CSWS shifted back to its maximum modal state, with the Labrador Current weakening and the frontal boundary of the LSSW retreating northeastward along the SS. As the supply of LSSW to the region decreased, ATSW returned to its previous position adjacent to the shelf break. By the end of 1999, incursions of warmer, more saline ATSW into the region’s shelf waters, combined with local warming, returned hydrographic conditions to a state resembling that prior to the modal shift of the CSWS triggered by the 1996 drop in the NAO Index.
Although the NAO-associated changes in slope water circulation have been relatively well documented from the SS to GB, the same cannot be said for the freshening of shelf waters observed from Newfoundland to the MAB during the decade from the late 1980’s to the late 1990’s (Smith et al., 2001; Drinkwater et al., 2002; Mountain 2002; 2003). While the source of the low-salinity water responsible for this freshening is upstream of the SS, the relative contributions from shelf sources in Newfoundland or further north versus the Gulf of St. Lawrence have not been fully resolved (Drinkwater et al., 2002; Frank, 2003). In the proposed research, we will focus our working group’s attention on variability in the salinity and volume of shelf waters supplied to the SS, GOM, GB, and MAB from upstream sources. There are two hypotheses associated with the observed freshening of shelf waters that we plan to investigate. The first focuses on inter-decadal variability, hypothesizing that the quasi-decadal salinity anomalies in the Labrador Sea, including the Great Salinity Anomaly of the 1970’s (Belkin et al., 1998) and the comparably strong salinity anomaly of the 1990’s (Hakkinen, 2002), enhance the supply of cooler and fresher Scotian Shelf Water (SSW) to the GOM, which, in turn, leads to the decadal-scale freshening and larger volume of Middle Atlantic Bight Shelf Water (MABSW) observed downstream (Mountain 2002; 2003). The second focuses on inter-annual variability, hypothesizing that phase shifts in the NAO affect the volumes and hydrographic properties of both SSW and slope water supplied to the GOM, which, in turn, leads to the year-to-year hydrographic and volume differences in the MABSW observed downstream. Determining the validity of both hypotheses will be important as we examine the implications of this lower salinity water on vertical stratification and production processes in the GOM and on GB. It will also be important as we extend our ecosystem studies to the MAB since the supply of MABSW formed in the GOM influences the position of the MAB shelf-slope front and the volume of environmentally suitable habitat on the shelf for cooler water species at the southern limits of their range.
Hydrographic
data for the time-series analyses used to explore the above hypotheses will be
drawn from the databases maintained by the NOAA Oceanographic Data Center,
Bedford Institute of Oceanography (BIO), and
We
anticipate using the results from these analyses of hydrographic and satellite
data to provide the foundation for future modeling and observational studies.
Dr. Hakkinen currently runs an Arctic-North Atlantic circulation model, based
on the Princeton Ocean Model, which is forced by appending atmospheric
anomalies from the NCEP/NCAR Reanalysis to a monthly climatology (Hakkinen,
1999). The model has been shown to simulate the salinity anomalies
characteristic of the 1970s and 1990s, reproducing the effects of southward
current anomalies and reduced meridional overturning on the downstream
hydrography (Hakkinen, 2002). Our research team will investigate the
feasibility of coupling such climate-forced, basin-scale circulation models
with the kind of regional-scale, physical-biological models currently used by
US GLOBEC investigators (reviewed by Hofmann and Lascara, 1998). This coupling
of basin- and regional-scale models can only achieve its full potential if it
is accompanied by the collection and analysis of new observational data,
especially the hydrographic data collected in Canadian waters by the Department
of Fisheries and Oceans (DFO)
Hypothesis 2: Remote forcing from the
The NAO-associated changes in ocean circulation patterns observed over the past 40 years have had a profound impact on marine ecosystems from the SS to GB (Greene and Pershing, 2000; Conversi et al., 2001; MERCINA, 2001; 2003; 2004; Piontkovski and Hameed, 2002; Drinkwater et al., 2003; Pershing et al., 2004; Sameoto, 2001; 2004). The springtime zooplankton biomass and secondary production in this region is dominated by the copepod species Calanus finmarchicus. An annual C. finmarchicus Abundance Index, derived from continuous plankton recorder (CPR) surveys conducted in the GOM, provides a good indicator of changes in the modal state of the CSWS (Fig. 2B, C) (Greene and Pershing, 2000; MERCINA, 2001; 2003). During the decade of the 1960’s, when the NAO Index was predominantly negative, and the CSWS was in its minimum modal state, slope water temperatures and C. finmarchicus abundance were relatively low. During the 1980’s, when the NAO Index was predominantly positive and the CSWS was predominantly in its maximum modal state, slope water temperatures and C. finmarchicus abundance were relatively high. During each of the maximum- to minimum-modal shifts in the CSWS after 1980, C. finmarchicus abundance declined in subsequent years. The modal shift during 1981-83 preceded a large, single-year decline in abundance during 1983. The modal shift during 1988-91 preceded a large decline in abundance that persisted throughout the early 1990’s. Then, after C. finmarchicus abundance began building up again during the mid-1990’s, the NAO Index underwent its drop of the century in 1996. This event triggered the intense modal shift of the CSWS during 1997, which, in turn, led to very low abundances of C. finmarchicus during 1998 and early 1999. The mechanisms underlying these climate-driven changes in C. finmarchicus abundance have not been fully resolved; however, they appear to be linked to the advective supply of this species into the GOM/SS region from the slope waters (Greene and Pershing, 2000; MERCINA, 2001; 2003; 2004).
Marine
ecosystem responses to NAO-associated oceanographic changes also have been
detected at trophic levels both lower and higher than the one occupied by C. finmarchicus. A time series of the Spring
Phytoplankton Color Index, a qualitative measure of phytoplankton standing
stock, was analyzed by Pershing (2001) using the same CPR survey dataset as the
one used to derive the C. finmarchicus
Abundance Index. He was able to
show that this admittedly crude index of standing stock exhibits many features
in common with the C. finmarchicus
Abundance Index, including low values in the 1960’s and high values in
the 1980’s. Pershing (2001)
hypothesized that these changes in the Spring Phytoplankton Color Index reflect
modal shifts in the CSWS and nutrient limitation of phytoplankton
production. This hypothesis is
consistent with differences in nutrient concentrations associated with the two
slope water types, as ATSW is characterized by high concentrations of nitrate
and silica, and LSSW is characterized by low concentrations of these nutrients
(Petrie and Yeats, 2000). It appears that the elevated nutrient concentrations
in ATSW are the result of Gulf Stream cross-frontal exchange processes bringing
nutrients to the
With regard to higher trophic
levels, we have developed a stochastic model showing that most of the
variability in right whale calving rates can be explained by NAO-associated
fluctuations in C. finmarchicus
abundance (Fig.
2, 3) (Greene et al., 2003; Greene and Pershing, 2004). The calving rates predicted by
this model capture the overall patterns very well, especially the large
fluctuations of recent years. Both multi-year declines observed during the
early 1990’s were reproduced by the model. In addition, it accurately predicted the
dramatic increase in right whale calves born during 2001. These results have
given us confidence in the model’s predictive capability, not only to
hindcast past events, but also to forecast right whale reproductive performance
at least one year into the future.
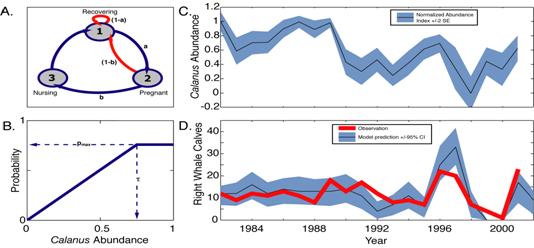
Figure 3.
Right whale reproduction model. A. Diagram of reproductive cycle, with
transitional probabilities between states indicated. A whale in any of the
three states, pregnant, nursing, or recovering, will move to the next state
with a probability determined by Calanus finmarchicus abundance in that
year. B. The transitional
probabilities are simple functions of Calanus finmarchicus abundance as
described by the parameters, t, the
saturating food level, and pmax, the maximum transitional
probability. C. The Calanus finmarchicus Abundance Index as determined from
Continuous Plankton Recorder surveys in the
As we look to expand the scope of our research beyond the NAO and beyond the SS/GOM/GB region, we will once again turn our attention to the analysis of existing time-series data sets. A recent principal components analysis of CPR zooplankton time-series data from the GOM has shown that several copepod species (Centropages typicus, Oithona sp., Pseudocalanus spp., and Metridia lucens) exhibit a mode of variability in abundance that is distinctly different from the one exhibited by C. finmarchicus (Pershing et al., 2005). In contrast to C. finmarchicus, these species increased dramatically in abundance from the late 1980’s until a rapid decline occurred in 2002. Pershing et al. (2005) hypothesized that the assemblage of copepod species exhibiting this mode of variability may have been responding to the decade-long freshening of the Northwest Atlantic shelf described previously, with its associated enhancement of winter-time stratification and primary production (Durbin et al., 2003).
We propose
to follow up on these earlier studies by conducting comparable principal
components analyses of zooplankton time-series data from the MAB to
In addition to analyzing zooplankton data, we propose to analyze Phytoplankton Color Index data from the New York-Bermuda transect CPR surveys and chlorophyll fluorescence data from the shelf-wide research vessel surveys. We also intend to analyze satellite ocean color data from the Sea-viewing Wide Field-of-view Sensor (SeaWiFS) mission. Dr. Bruce Monger and several NASA Space Grant-supported students at Cornell have conducted comparable studies in the SS/GOM/GB region since the launch of the SeaWiFS sensor in 1997, and the proposed effort simply expands the domain of these studies to include the MAB. Ms. Yianna Samuel, a graduate student at Cornell, will assist Dr. Monger with the expanded effort. We have found evidence in the SS/GOM/GB region that phytoplankton production and the seasonal cycle may be altered by NAO-associated changes in nutrient supply and salinity-anomaly-induced changes in the timing of stratification (Pershing, 2001; MERCINA, 2004). We hypothesize that analyses of in situ and satellite data will reveal comparable responses of phytoplankton primary production to climate-associated changes in hydrography and circulation in the MAB.
D. Broader Impacts
Climate and Ecosystem-Based Resource
Management
A
major goal of our project is to develop a predictive understanding of climate
impacts on marine ecosystems in the
Recently,
two members of our research team (CHG and AJP) submitted a proposal to the
In making the leap from documenting climate-associated ecosystem responses to forecasting the future of managed populations, care must be taken to account for the ecological mechanisms linking the demography of these species to the ecosystem. For exploited fish populations, single-species fisheries management typically has involved setting catch limits based on environmentally invariant stock-recruitment models. In contrast, ecosystem-based fisheries management requires the development of models that can predict recruitment from stock assessment data combined with data on environmental variability.
To illustrate one method for incorporating climate-associated environmental variability into fisheries recruitment models, we briefly describe two recent studies by Brodziak et al. (2005) and Pershing et al. (2005). These authors, which include invited participants to our proposed NCEAS WG, used the following standardized recruitment model to distinguish between factors internal to the population and those arising from external environmental forcing:
![]() (1)
(1)
where Rs is the standardized recruit per
spawner index, R and S are the recruitment and spawning stock
biomass inferred from observations (subscript obs) and the virtual population assessment (subscript VPA), respectively, and ![]() is the standard deviation of the
observed-VPA series. The standardized recruitment model subtracts the component
of recruitment due to internal population factors from the observed
recruitment, leaving as a remainder the component due to external environmental
forcing.
is the standard deviation of the
observed-VPA series. The standardized recruitment model subtracts the component
of recruitment due to internal population factors from the observed
recruitment, leaving as a remainder the component due to external environmental
forcing.
Brodziak et al. (2005) showed that time series of Rs for eight of 12 commercially important fish stocks investigated in the GOM/GB region exhibited significant, time-lagged cross correlations with the NAO Index time series. Their preliminary results suggest that the external, environmentally forced recruitment of these stocks may be linked to physical and/or biological processes responding to NAO forcing.
Pershing et al. (2005) took a different approach to the problem and drew conclusions that, at first, appear to be somewhat at odds with those of Brodziak et al. (2005). First, these authors used principal components analysis to establish two distinct modes of zooplankton variability (the “C. finmarchicus” mode, the “other copepods” mode) and related these to the two climate-associated changes in regional physical oceanography described previously (the NAO-forcing of the CSWS and the 1990’s decadal freshening from upstream). They then conducted cross-correlation analyses between time series of the zooplankton modes of variability and time series of Rs for the same 12 fish stocks studied by Brodziak et al. (2005). Results from the analyses conducted by Pershing et al. (2005) indicate that the decadal freshening of the region during the 1990’s had a greater impact on more copepod species and fish stocks than the changes brought about by NAO forcing. The results from these two studies are compatible when one recognizes that zooplankton need not be mediating the statistical associations between NAO forcing and fish recruitment reported by Brodziak et al. (2005). For fish stocks, like GB haddock, in which the time series of Rs is positively cross correlated with time series of both the NAO Index and the mode of zooplankton variability linked to NAO forcing, the results might encourage further investigation of hypotheses linking C. finmarchicus abundance to GB haddock recruitment success. For the other fish stocks, with time series positively cross-correlated with NAO Index but not with the “C. finmarchicus” mode, it may be more productive to explore other types of hypotheses first. The purpose of highlighting these two studies here is to illustrate the kind of insights and synergy that can emerge by assembling WGs composed of oceanographers, fisheries scientists, and fisheries managers. The broader perspective of such multidisciplinary WGs frequently leads to the formulation of more thought-provoking hypotheses and eventually a deeper understanding of the processes regulating fish abundance in the sea.


Educational Impacts
A significant portion of the funds requested in this proposal will be used to support the training of two graduate students, Ms. Louise McGarry and Ms. Yianna Samuel. Ms. McGarry participated as a research technician in all of our GLOBEC NWA autumn cruises to the GOM during the late 1990’s. She also participated in several GLOBEC NWA scientific investigator meetings during and subsequent to the field program. The relatively large allocation of the requested funds to support graduate students is consistent with our commitment to integrating education with research. During the past decade, in addition to the educational outcomes cited in Results from Prior NSF Support, two members of our research team (CHG and BCM) have run advanced courses in bioacoustical and satellite oceanography that have trained more than 150 students from greater than 20 countries. Female graduate students consistently have been given prominent leadership roles in these courses. The support requested here for Ms. McGarry and Ms. Samuel, teaching assistants in three of these courses since starting graduate school, will enable us to continue providing them with exceptional learning opportunities during the remainder of their graduate educations.
In addition to graduate education, we will continue to involve undergraduates in our research, just like the seven supervised during our previous US GLOBEC studies.
E. Project Organization and Time Table
Drs. Charles Greene and Andrew Pershing will serve as the project’s principal investigators, sharing in the responsibilities of organizing working group meetings, overseeing the research team’s activities, and supervising the graduate research assistants. Drs. Hakkinen, Monger, and Mountain will serve as senior personnel on the research team, overseeing the research issues outlined on p. 8 of the proposal. Drs. Loder, Head, Smith, and Sameoto also will serve as participants on the research team and have responsibility for several of the research issues outlined on p. 8 and 10 of the proposal. The remaining working group members (see Table 1) will participate in the regional meetings (see Table 3) where they will work with the research team to steer the research agenda and assist in the interpretation of results.
Table 3.
Time Table
ACTVITY TOPICS _____________ TIME_______
First
Regional WG Meeting Autumn
2005
-
Review Previous Work Related
to Hypotheses One and Two
-
Assignment of Research Tasks
-
Plan AGU/ASLO Special Session Talks
First
NCEAS WG Meeting Autumn
2005
-
Review of Analytical Methods
-
Identification of New Data Sets
-
Assignment of Research Tasks
-
Plan AGU/ASLO Special Session Talks
Data
Analyses
Autumn 2005 – Summer 2006
Proposed
Special Session: AGU/ASLO Ocean Sciences Meeting Winter 2006
Second
Regional WG Meeting Autumn
2006
-
Reports on Research Accomplishments
-
Planning Publications/Other Products
-
Assignment of New Research Tasks
-
Plan AAAS Special Session Talks
Second
NCEAS WG Meeting Autumn
2006
-
Reports on Research Accomplishments
-
Planning Publications/Operational Products
-
Assignment of New Research Tasks
-
Plan AAAS Special Session Talks
Special
Session: AAAS Annual Meeting Winter 2007
Data
Analyses, Manuscript Preparation Autumn 2006 – Summer 2007
Combined
Regional and NCEAS WG Meeting Autumn
2007
-
Reports on Research Accomplishments
-
Complete Publications/Operational Products