Session IV: Hot Topics Discussion
Steve Bollens, Chair
Greg Lough, Rapporteur
Session Summary
The "Hot Topics" Session was meant to serve as a forum for investigators to briefly (5 minutes) present novel and intriguing research results that either did not fit readily under the theme areas established for the other three sessions or for which investigators did not have adequate time to present during the other sessions. As such, some of these presentations addressed topics only indirectly related to the GLOBEC target species (sand storms on Georges Bank by P. Wiebe; the Chaetoceros patch by D. Gifford et al.; planktonic hydroid growth by Bollens et al.), whereas others addressed more specific aspects of the population biology of the target species (Calanus mating behavior by C. Miller; inter-decadal zooplankton abundances by Landsteiner et al.; Calanus egg viability by J. Runge). But throughout all of this session's diverse presentations runs the common thread of trying to elucidate the intricate web of biological-physical interactions that comprise the Georges Bank ecosystem.
Mate Finding Behavior in Calanus marshallae
Direct observations and video recordings of mate finding behavior in Calanus marshallae revealed general patterns of female and male activity which lead to encounters and transfer of spermatophores. These patterns were outlined in the talk and will be published in the near future.
Sand Storms on Georges Bank
During the July 1995 broad-scale cruise on Georges Bank (Albatross IV 9508 Figure 1), the towed high frequency (420 kHz) acoustics profiling system encountered extremely high acoustical volume backscattering in the shallower regions of the Bank. One section from the middle of the southern flank (Station 9) to the crest of the Bank (Station 11) and then along the crest to the east (Station 12), illustrated the phenomenon particularly well (Figure 2). In waters that were stratified (depths of ~65 to ~50 m), the acoustical record showed strong surface and subsurface horizontal layers associated with the mixed layer and pcynocline. In a region where the bottom shoaled abruptly from 50 to 40 m, the water column became well mixed and the volume backscattering became dominated by fine-scale vertical lineations from the surface to the bottom. The intensity of the backscattering increased substantially as the depth shoaled to the bank crest. Along the top of the crest, patches of very high volume backscattering were observed which extended from the top to bottom. We suggested previously that such vertical lineation structure is due to the interaction of the particle field with secondary cell circulations associated with strong tidal flow over a rough bottom (Wiebe et al, in press).
On this occasion, net tows (with 150 um mesh nets) and pump sampling (with ~50 um mesh netting) done at stations along the crest contained substantial quantities of sand, even at the surface. An experiment conducted later in the cruise at stations 36, 37, and 38 were specifically designed to examine the relationship between volume backscattering and sand in the water column as determined from pump samples.
Four hypotheses have resulted from this finding:
The Chaetoceros socialis Patch
C. socialis colonies were present in densities of ~5 colonies ml-1. Colonies were extremely fragile and were observed only in water siphoned gently through wide-bore tubing from the top of Go-flo bottles. Water drained through spigots at the bottom of Go-flo bottles did not contain intact colonies. The colonies consisted of a few hundreds to a few thousands of cells arranged in chains wrapped into a spheroidal shape. Thin threads converged toward the colony center, where consortia of small heterotrophic flagellates were located. Individual colonies contained on average 300 pg chlorophyll a (range 120-490; n=45 colonies) and 800 pg phaeopigment. Grazing experiments indicated that the colonies were consumed by stage C6 female Calanus finmarchicus.
We suggest that the patch represents a potentially extensive (in space and time) 3-dimensional structure in the water column on the southern flank of Georges Bank. As such, it may provide a concentrated nutritional resource to consumer organisms, or may function as a specific habitat for the nano- and microplankton organisms associated with it.
Inter-Decadal Comparison of Monthly Mean Zooplankton Abundances on Georges Bank
We examine data collected during 1939-41 (R/V Atlantis) and 1977-87 (MARMAP) for any possible inter-decadal changes in zooplankton abundances. Currently the Atlantis data and the MARMAP data allow the only quantitative inter-decadal comparison of zooplankton abundances on Georges Bank. The Atlantis data, a largely unpublished and recently uncovered data set, consist of vertically-stratified net samples collected at 21 to 52 stations on each of 11 cruises using a Clarke
Bumpus plankton sampler (12.7 diameter mouth opening; ~250 mm mesh netting). The MARMAP data consist of vertically-integrated net samples collected at 15 to 50 stations on cruises conducted 6 to 12 times per year using a bongo net (61 cm diameter mouth opening; 333 mm mesh netting). Only adult stages for selected species were compared so as to avoid possible biases due to the different mesh sizes. Differential avoidance of the two samplers may be another possible sampling bias; however, work by Clarke et al. (1943) showed for Sagitta elegans that this is unlikely to be a problem.
We present a preliminary comparison of the monthly mean abundances of the dominant zooplankton species (Calanus finmarchicus, Pseudocalanus minutus, Metridia lucens, and Centropages hamatus) on the Bank (< 200 m) (Figure 1). Although seasonal abundance patterns for C. finmarchicus, P. minutus, and C. hamatus are comparable between the two time periods, absolute abundances of C. finmarchicus, M. lucens and C. hamatus are generally less during 1939-41 than 1977-87. This could be due to any number of biological or physical processes during this period. One intriguing possibility is inter-decadal variability in the abundance of vertebrate predators. For example, abundances of mackerel, a known predator of C. finmarchicus (Madin et al.; Session II), were relatively low during the 1970s (Fogarty et al.; Session II).
Future work with the MARMAP data will examine the effect of storminess on mean abundances and spatial distributions of zooplankton and the results will be compared with our investigations of the Atlantis data (Bollens et al.; Session I).
Reference:
Clarke, G. L., E. L. Pierce, and D. F. Bumpus. 1943. The distribution and reproduction of Sagitta elegans on Georges Bank in relation to hydrographical conditions. Biol. Bull. 85: 201
226.
Figure 1: Monthly mean abundances and variances [vertical bars = 2
SE] of adult zooplankton on Georges Bank (bottom depths < 200 m);
longitude east of 69.1W). [circles = Atlantis (1939-41);
solid diamonds = MARMAP (1977-87)]
Influence of Diatoms on the Viability of Early Life Stages of Copepods: Implications for Understanding Variability in Copepod Recruitment Rates on Georges Bank
Recent studies by S. Poulet, A. Ianora and co-workers show that ingestion of certain diatom species by female copepods inhibits development of their offspring. Experimental results are most consistent with the hypothesis that these diatoms contain a toxin that either blocks embryogenesis entirely or induces deformities in the early naupliar stages. In laboratory experiments, we have observed that feeding by Calanus finmarchicus females on the common diatom, Thalassiosira nordenskioldii, greatly reduces hatching success of their eggs. A control group of females feeding on the dinoflagellate, Prorocentrum micans, produced eggs that hatched with a 90% success rate. During 1995 process cruises on Georges Bank, eggs released by C. finmarchicus typically hatched with a success rate of 60-90%, although at some stations only 30-35% of the C. finmarchicus eggs were viable
(Figure 1). These observations suggest that the influence of maternal diet on the viability of offspring may contribute to patterns of variability in recruitment into the region's copepod populations.
Figure 1. Hatching success of eggs released by C. finmarchicus females captured at Georges Bank stations during 1995 process cruises.
Growth of Planktonic Hydroids: Experimental Evidence for the Importance of Mixing, Particulate Food and DOM
During spring of 1994, we found huge numbers of hydroids (principally Clytia gracilis) suspended in the plankton on Georges Bank. We subsequently showed that the hydroids were capable of consuming cod larvae and young copepods; in the latter case having a potentially devastating effect on their prey (Madin et al., In Press; Session II). Based on these initial findings we mounted a much more extensive field study during spring and summer of 1995 on the distribution and abundance of hydroids in both the water column and on the bottom of Georges Bank (see Sullivan et al.; Session I). Additionally, we undertook a series of laboratory investigations on the feeding, growth and life history characteristics of planktonic hydroids, some preliminary results of which we summarize below.
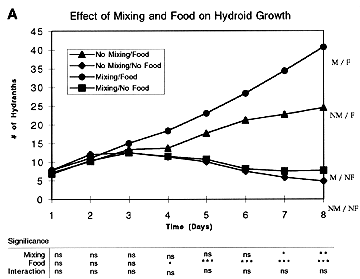
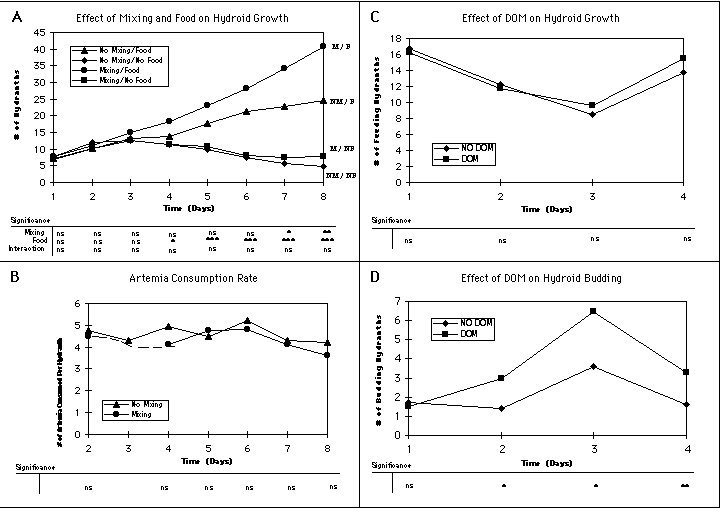
In the first experiment we established replicated (6) 1 liter jars in each of four treatments (Mixing + Food; Mixing + No Food; No Mixing + Food; No Mixing + No Food) in a 15° C cold room. The mixing treatments were bubbled at a rate of ca. 500 bubbles per minute using a ca. 1 mm bore glass tube in each jar. The food treatments were adjusted to 160 Artemia nauplii per liter at the start of each day. Filtered seawater was exchanged in all jars once each day. There was a clear effect of food on the growth of hydroid colonies, with the mean number of hydranths per colony diverging between the food and no food treatments after approximately 4 days
(Figure 1A). Perhaps more interesting, however, was the additional effect on hydroid growth due to mixing
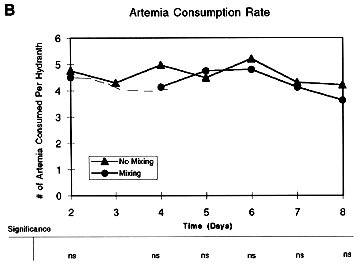
(Figure 1A). Although we initially thought this effect might be due to turbulence-enhanced feeding via greater predator-prey contact rates in the mixing treatments, a comparison of the consumption rates in the Mixing/Food versus No Mixing/Food treatments showed no significant differences between treatments
(Figure 1B), i.e., the enhanced growth rate observed could not have been due to greater ingestion in the mixing treatment.
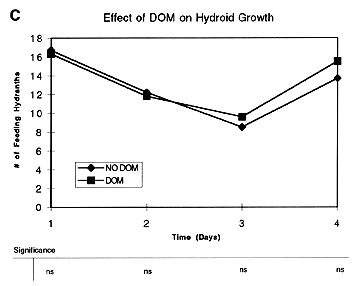
This led us to consider other possible advantages that mixing might confer on hydroid growth, including better oxygenation, waste removal and uptake of DOM. The former was discounted in that all experimental containers remained well oxygenated through-out the experiment. The possible role of DOM was tested by establishing two treatments of 10 replicates each: 1) a DOM treatment, consisting of filtered and autoclaved seawater with additions of 1 mM of glucose and 62.5 nM of 16 amino acids; and 2) a No DOM treatment consisting of filtered and autoclaved seawater only. After four days the results showed no significant differences in number of feeding hydranths between treatments
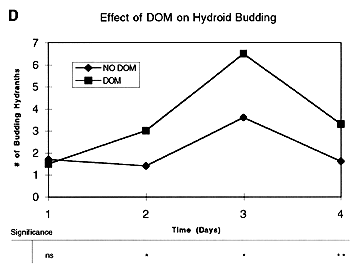
(Figure 1C), although the trend was clearly towards a divergence. A comparison of the number of budding hydranths, however, showed a statistically significant difference between treatments on days 2-4
(Figure 1D). At this point we can not distinguish between a direct effect of DOM uptake by the hydroids versus an indirect effect due to high bacterial growth and ingestion of bacteria by the hydroids. However, the not uncommon observation in other taxa of invertebrates showing direct DOM uptake suggests this as the likely mechanism for enhanced growth of the hydroid colonies in the DOM treatment.
Results of these laboratory experiments, when combined with those from the field studies, will allow us to determine which biological and physical processes are responsible for maintaining these abundant and important predators on the shoal region of Georges Bank.
Charles B. Miller1 and Atsushi Tsuda2
1Oregon State University
2University of Tokyo
Peter H. Wiebe
Woods Hole Oceanographic Institution
References:
Figure 2. A 3-D view of the volume backscattering at 120 kHz along tracklines between Standard Stations 9-10, 10
11 and 11-12. The first two tracklines run from south to north from the stratified region on the southern flank of the Bank to the top of the Bank. The latter runs from west to east (perpendicular) to the first two lines along the top of the Bank. Note the very intense backscattering along the top of the Bank which may be due to sand particles in the water column.
Dian J. Gifford1, Michael E. Sieracki2, Cabell S. Davis3 and Scott M. Gallager3
1University of Rhode Island
2Bigelow Laboratory for Ocean Science, CA
3Woods Hole Oceanographic Institution
A patch of colonial Chaetoceros socialis was observed on the southern flank of Georges Bank in June 1995. The patch was resolved by video and fluorometry sensors on the Video Plankton Recorder and by the profiling fluorometer on Endeavor's Neil Brown CTD package. While we were unable to map the patch in its entirety, the patch covered an area of at least 6 x 30 miles. It was located at approximately 20-30 meters depth, coincident with the pycnocline. We observed the same feature on the southern flank during US-GLOBEC cruise CI9407 in June, 1994. Bigelow (1924) described a similar feature in the same region during 4 months in the spring of 1920.
Mary Landsteiner1, Stephen Bollens1, Cabell Davis1, Andy Solow1 and Jack Green2
1Woods Hole Oceanographic Institution
2National Marine Fisheries Service, Narragansett
Jeff A. Runge1, M. Starr1 and S. Plourde2
1Institut Maurice Lamontagne, CA
2Université du Québec à Rimouski, QC
Steve Bollens, Larry Madin, Erich Horgan, Stephanie Concelman, Brenda Burkhalter
and Mari Butler
Woods Hole Oceanographic Institution
Figure 1. Results of experiments using particulate food (A and B) and DOM (C and D) to test for effects of various treatments on hydroid growth. Data points are means of 6-10 replicates for each treatment, with differences between means examined by ANOVA (see table below each plot; ns = non-significant; * = p < 0.05; ** = p < 0.01; *** = p < 0.001).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}