|
GLOBEC scientists study the coupling between physical and biological processes, using past and
present climate variability as a proxy for future climate change. The research approach (and challenge)
is to combine modeling, retrospective analysis of historical data, and process research into an
integrated program that will produce regional climate change scenarios and quantitative assessments of
the sensitivity of selected marine ecosystems to climate variability and climate change.
|
|
|
Recent studies have documented that the
physical and biological dynamics of the eastern Pacific are sensitive to natural climate variability on
time scales ranging from seasonal to interdecadal, and spatial scales from local to basin-wide.
Ecosystem structure is known to be closely coupled to variations in physical forcing, thus sensitivity
of the coupled physical-biological system to climate variability implies great sensitivity to climate
change.
|

|
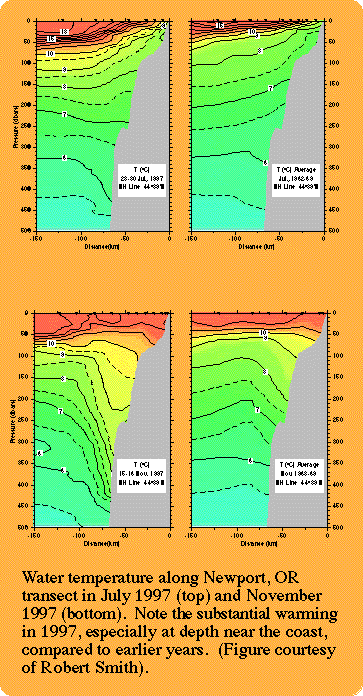
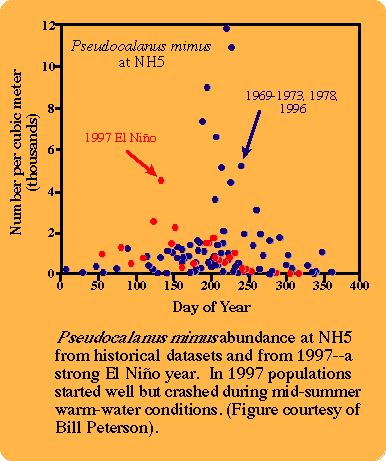
The physical structure, circulation dynamics and biology of the eastern North Pacific respond strongly to El Niño-Southern Oscillation (ENSO) and decadal time-scale regime (climate) forcings. As an example, sea surface temperature (SST) anomalies for coastal and offshore regions off the west coast of Central and North America show both the influence of El Niño events (warm water at 0-30°N during 1972-73 and at all latitudes during 1982-83), and regime shifts (warm anomalies during winter and spring north of 24°N since 1976-77) on nearshore and offshore SST.
|
|
|
The strong connections between the eastern North Pacific and basin-wide oceanic and atmospheric
circulations provide linkages to global scale climate changes, as shown by the well documented
response of the California Current System to the basin-wide, interannual ENSO variability (see papers
in Wooster and Fluharty, 1985; Chavez, 1996). The impact of ENSOs on marine populations is best
documented for regions south of 35°N (Point Conception, CA) in the eastern North Pacific (see e.g.,
Barber and Chavez, 1986; McGowan, 1985; Smith, 1985; Fiedler et al. 1986). Weak El Niño events
probably have little impact on marine populations north of the tropics in the eastern North Pacific,
whereas some (but not all) strong El Niño events, like that of 1982-83, may impact populations from
the equator to the subarctic (Miller et al. 1985; Pearcy et al. 1985). The average size of coho salmon captured in the Oregon fishery during the strong 1983 El Niño was the smallest on record, suggesting that low food availability led to poor growth (Pearcy 1992).
|
 Fisheries production domains and general circulation in the NEP Ocean (From Ware and McFarlane, 1989). |
The evidence cited above indicates that the 1983 El Niño had deleterious effects on the productivity of the pelagic environment off the Pacific Northwest, which negatively impacted commercial harvests of some fish stocks. It should be noted, however, that the biological impacts of El Niños can be positive or negative, and is often region and population specific. For instance, strong recruitment and good year classes of stocks of Pacific sardine and jack mackerel off California resulted from the 1958-59 El Niño, but Pacific Ocean perch, Dover sole and English sole off Oregon and Washington were negatively impacted by the 1958-59 El Niño (Bailey and Incze, 1985). The 1982-83 El Niño reduced the growth of northern anchovy juveniles and adults, but expanded the region in which spawning occurred (Fiedler et al. 1986). El Niño events can impact marine populations in a multitude of ways: 1) by altering food production, distribution, availability and phasing (timing) relative to the consumer populations; 2) altering transport regimes and residence times; 3) environmental warming that alters physiology or causes range shifts; and, 4) altering the intensity of predation pressure by introducing new or changed abundances of predators to a region (i.e., range expansions).
|
El Niño's, with time-scales of ca. 2-7 years, are perhaps the most noticeable temporal
variability in the NEP, but there are substantial longer-term variations that impact marine resources as well. The climate of the subarctic North Pacific Ocean changed during the late 1970s (Fig. 2). The Aleutian
Low intensified during winter (Trenberth and Hurrell 1994) and coastal sea surface temperatures rose
rapidly by several degrees (Rogers and Ruggerone 1993; Royer 1989; Graham 1995). The deepening
of the Aleutian Low resulted in more vigorous cyclonic circulation of the North Pacific subarctic gyre,
and a deepening of the mixed layer in the North Pacific subtropical anticyclonic gyre. A dramatic shift
in ocean climate also occurred in the California Current System. The warm phase/cool phase shifts in
the California Current appear to be linked to the intensity of the Aleutian Low (Trenberth 1990;
Graham 1994; Miller et al. 1994b; Trenberth and Hurrell 1994). An earlier shift from
the warm phase to the cool phase occurred in the mid-1940s (Miller et al. 1994a). The shift in
1976-77 is particularly well documented because data on atmospheric, sea surface and subsurface
ocean conditions were sufficient to demonstrate the basin scale nature of the shift.
|
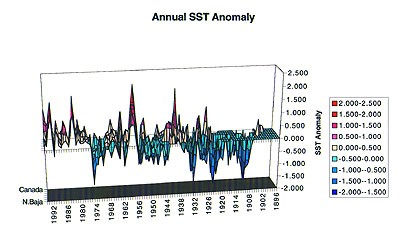 Annual sea-surface temperature anomalies from coastal records spanning from Northern Baja to Canada and for the period 1896-1996. Note that time proceeds from right to left. Large El Niño events and the more recent "regime shift" to warmer conditions are evident in the record. (Figure Courtesy of Franklin Schwing). |
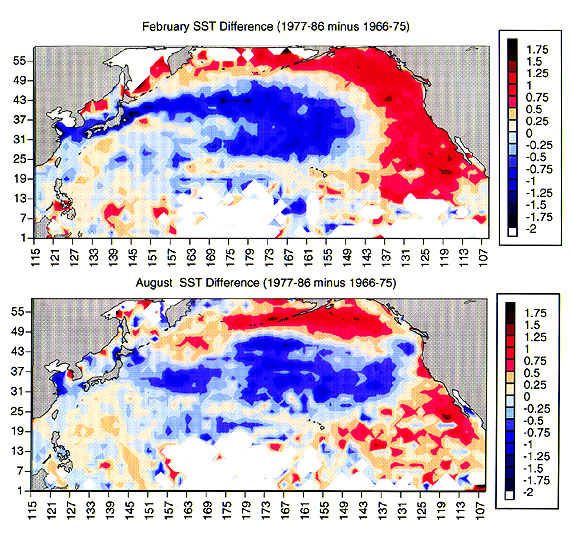
February (top) and August (bottom) SST differences between 1977-86 and 1966-75. Large positive changes (warmer water in more recent regime are red; cooler water is blue) are widespread along the west coast during winter and summer, with largest differences during winter. (Figure Courtesy of Franklin Schwing).
Polovina and coauthors (Polovina et al. 1994; 1995) document the changes in North Pacific winter and spring mixed layer depth (MLD) and mixed layer temperatures (MLT) of the North Pacific. MLD in the subtropical and transition zones of the North Pacific were 30-80% deeper during 1977-88 than during 1960-76. Across the subarctic zone, including the Gulf of Alaska, MLD was 10-30% shallower during the later period than during the earlier. Using a plankton production model, Polovina et al. (1995) argue that the changes in winter and spring MLD and MLT could have resulted in 50% higher primary and secondary production in both the subarctic and subtropical realms of the North Pacific. They conclude that decadal time-scale and basin spatial-scale changes in MLD and MLT in the North Pacific are related to the intensification of the Aleutian Low Pressure System, and provide a mechanistic link between atmospheric circulation variability and oceanic ecosystem productivity. Productivity of higher trophic levels (lobsters, birds, seals) off Hawaii appears to respond also to the change in ocean climate of the late 1970's (Polovina et al. 1994).
Biological responses to interdecadal basin (or global) scale variability have been recognized in both the California Current System (CCS) and the Coastal Gulf of Alaska (CGOA). Perhaps the best documented are in the abundance (or distribution) of fish populations, like the salmonids (see collection of papers in Beamish 1995 and Beamish and McFarlane, 1989; Beamish 1993; Bailey et al. 1995; Hare and Francis 1995; Francis and Hare 1994; Brodeur and Ware, 1995) and small pelagic fish (sardines and anchovies) (Lluch et al. 1989; Kawasaki 1992). One cannot help but be impressed by the remarkably similar patterns in abundance (as indicated by catch) of Pacific salmon from the subarctic Pacific, and sardines from the California, Peru/Chile and Japanese fisheries over the past century. Periods of high and low catches of these stocks are related to northern hemisphere temperatures, with highest catches occuring during warm periods and vice versa (Klyashtorin and Smirnov, 1995). For the fisheries (figure below) for which records exist to the early 1900s, catches increased during the early part of the century, until about the mid to late 1930's. During the 1940s, each stock began to decline with lowest abundances in the late 1960s-early 1970s. Salmon catches rebounded sharply in the late 1970s and 1980s (Pearcy 1992). The commercial salmon harvest in Alaska in the 1980s exceeded that of the 1930s peak; however, some of this recent resurgence is due to salmon ranching (returns of hatchery released fish).
|
The catch records of sardines from the Pacific mirror that of the Alaskan salmon harvest. Historical
maximum catches of California and Japanese sardines in the 1930s-early 1940's were followed by a
rapid, precipitous decline during the succeeding three decades. Since the late 1970s, sardines in
Peru/Chile and Japan have rebounded strongly; in fact the maximum catch of Japanese
sardines during this latter period exceeded by a factor of two the maximum catches from the 1930s.
Commercial fisheries off Peru and Chile began only in 1960, when a fishery developed on the
abundant anchoveta; sardines at that time were relatively rare, but increased in abundance during the
1970s and 1980s. Unlike the case of the North Pacific salmon, there have been no remedial efforts
(hatcheries, habitat modification, etc.) involved in the sardine's recovery in these ecosystems.
California sardines have been increasing in abundance (with high population growth rate) since the late
1970s (Barnes et al. 1992), but the reestablishment of large stocks after the ocean conditions changed
has been slow because the stock was so depleted during the 1960s. The dramatic fluctuations in catches described above are likely due in part to changing patterns of fishing effort and the consequences of overexploitation. Nonetheless, the concordance among species suggests the effects of an overriding climate influence.
|
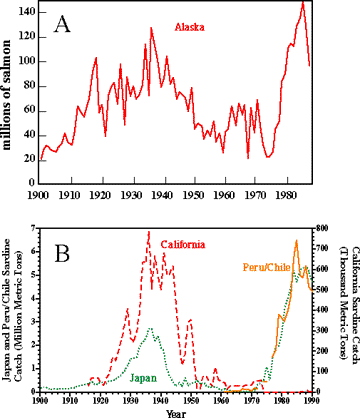 Commercial salmon harvest of Alaska, 1900-1988 (top). Data are from the Alaska Dept. of Fish and Game. (Redrafted from Pearcy, 1992). Historical catches in the sardine fishery (bottom) of Japan, California, and Peru-Chile. (Modified from Kawasaki, 1992). Note different ordinate scales. |
Long term, interdecadal scale variations in zooplankton biomass in the Southern California Bight (SCB) of the CCS are documented by Roemmich and McGowan (1994) using the CalCOFI data set. Zooplankton biomass in the SCB began a ca. 5-fold decline at approximately the time of the mid-1970's warming event in the CCS. Elsewhere, zooplankton biomass in the subarctic Pacific during the 1980s is ca. double that of 1956-1962, and the peak biomasses in the recent period are more coastally distributed than in the earlier period (Brodeur and Ware 1992; Brodeur et al., in press). These results suggest major shifts in the productivity of the subarctic gyre (increased) and the southern part of the California Current (decreased) during the present warm regime (warm marginal currents post 1976). We note that changes in the zooplankton populations in the northern part of the California Current during this period are not known because there was no systematic sampling (like CalCOFI) in this region.
|
To address questions about the physical and biological impacts of climate change requires data
spanning long time horizons--from the past, present and future. Each of these is a component of U.S.
GLOBEC studies: variability and change in the past is examined through Retrospective Data Analysis;
conditions and biophysical interactions at present are examined through Process-Oriented Field
Studies; and, documenting future variability and change is the rationale for instituting frequent,
sustained Monitoring of the environment. Modeling and Synthesis activities will integrate the results
from U.S. GLOBEC's studies, and from the research, monitoring, and retrospective activities, so that
the consequences of climate change on the coastal marine ecosystem can be evaluated and projected.
|
|
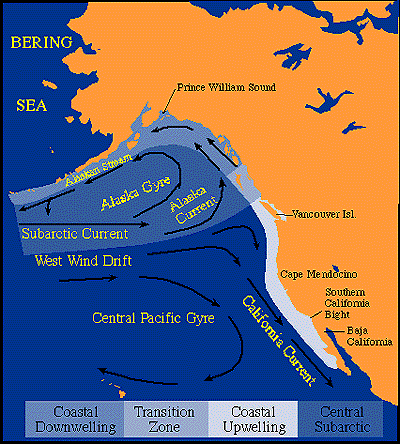
U.S. GLOBEC research in the Northeast Pacific will focus on the boundary currents--the equatorward flowing California Current along the southern region, the poleward and westward flowing Alaskan Current and Alaska Coastal Current in the north, and the interior west wind drift which diverges at the coast of North America and contributes to both boundary currents. Retrospective studies can utilize a rich history of time-series including research surveys of zooplankton and fish, commercial fisheries data, environmental data, and paleo records from sediments. These data sources can be used to define better the relation between large-scale climatic shifts, changes in population distributions and abundances, regional-scale transports, and local-scale processes, such as predation, upwelling/downwelling, mixed-layer depth, and zooplankton and fish production. Monitoring transects should be (re)established to provide ongoing time series of key processes at several locations off northern/central California, Oregon/Washington, and in the CGOA--preferably at locations with good historical records.
|
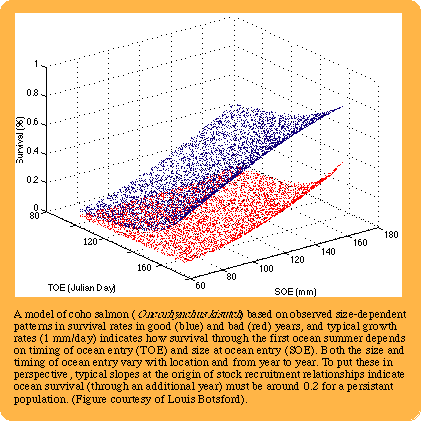
U.S. GLOBEC's Northeast Pacific process studies will investigate the way in which zooplankton and
juvenile salmon are affected by mesoscale circulation in the coastal ocean.
|
|
Mesoscale dynamics are a focus in the CCS and CGOA because they dominate much of the physical and biological dynamics (e.g., see satellite images), and because they differ regionally as a result of differences in wind stress, intensity of coastal upwelling/downwelling, coastal morphology, freshwater inflow, and the influence of advection, turbulence and buoyancy. Since climate-mediated changes in large scale atmospheric and oceanic forcing have substantial impact on mesoscale dynamics, a field research effort focused on this spatial scale is warranted. In addition to these physical differences along the west coast, there are regional differences in planktonic, benthic and fish assemblages, overall productivity, and the timing of production cycles. The northern domain--approximately from the Queen Charlotte Islands, BC to the Aleutian archipelago--of the Pacific coast is strongly influenced by coastal freshwater input, which results in an intense baroclinic, buoyancy-driven current along the coast, with eddies induced by the shear and topographic influences. Nursery areas and migration routes of both juvenile and adult fish parallel the resulting frontal boundary; the coastal current may be both a "conduit" for alongshore transport and migration and a "barrier" to cross-shelf motion (Thomson et al. 1989). The coastal Gulf of Alaska is a predominantly downwelling system. Conversely, southern British Columbia, Washington, Oregon, and northern and central California are predominantly upwelling systems. As described in earlier GLOBEC reports No. 7 (1991) and No. 11 (1994), Washington and Oregon north of Cape Blanco are characterized by winds that reverse seasonally, moderately strong upwelling (occuring in summer only), a linear coastline with few large promontories, predominantly alongshore advection, and few mesoscale features (e.g., eddies, filaments, offshore extending squirts, jets). This contrasts with the situation in northern and central California (to Pt. Conception in the south), where winds remain mostly upwelling favorable throughout the year, strongest in spring and summer, there are major promontories jutting into the ocean, offshore advection is large, and complex mesoscale features (squirts, jets and eddies) are prevalent. These regional differences in physical-biological linkages and the physical forcings provide a natural laboratory for comparing potential changes in marine populations due to climate variability and climate change over the larger, basin-scale.
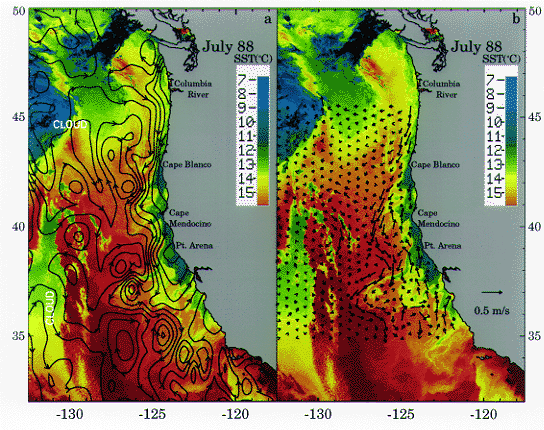
Summer (1988) circulation of the California Current. a) Geosat height from a single cycle in July 1988, contoured and overlaid on a coincident SST field; b) estimated surface velocity from automated feature tracking using five pairs of SST images over a 36 hour period, overlaid on SST. (From Strub and James, 1995).
| U.S. GLOBEC studies in the NEP will occur over an approximately 8 year period, with year 1 of the study being 1998. A timeline for the project is shown to the right. |
|
|
U.S. GLOBEC's Northeast Pacific process studies will focus on several target species
as shown at right.
|

|
This page was last updated on August 17, 2000.
Maintained by:
Hal Batchelder
COAS
Oregon State University
Corvallis, OR 97331-5503
phone: 541-737-4500; FAX 541-737-2064