The session had 29 presentations that addressed Bank-wide structure of the physics and biology as observed in recent field studies, in historic data sets and in modeling efforts. From the presentations two issues were identified for discussion. Winter time observations in the program found Calanus finmarchicus actively involved in growth and reproduction in the near surface layer. This finding diverged from the conventional Calanus life cycle paradigm, which would have the organisms in diapause at depth in the western Gulf of Maine in winter. Charlie Miller summarized the knowledge of the Calanus life cycle and led a discussion on the implications of the recent findings. The second issue identified was how to evaluate the relative importance of predation by different predator groups and the combined importance of predation compared to other factors in the life history of the target species. Michael Fogarty led a discussion of this issue. Summaries of both issues are included with the abstracts for the presentations in this session.
Large-Scale Hydrographic and Circulation Variability in the Georges Bank Region
John Loder, D.A. Greenberg and C.G. Hannah
Bedford Institute of Oceanography, CA
Implications for Georges Bank of hydrographic and circulation variability in the Gulf of Maine and Scotian Shelf regions are discussed, using the historical temperature and salinity database and a diagnostic circulation model (Lynch et al. 1992).
Model solutions indicate that, in contrast to outer Scotian Shelf banks, Georges and Browns Banks are located in the equatorward evolving barotropic pressure field arising from along-shelf wind stress, resulting in a significant barotropic as well as Ekman-layer current response. Topographic channelling results in most of the cross-shelf Ekman transport on the Scotian Shelf being compensated by barotropic flow through Northeast Channel and along the continental slope off Georges Bank. At storm-band periods (2-5d), the Northeast Channel flow excites a topographic wave along the southern Georges slope. These and other aspects of the barotropic response to wind stress are described in Greenberg et al. (1995).
The climatological seasonal variations in hydrography and baroclinic circulation are examined using 3-d seasonal-mean temperature, salinity and density fields obtained from the historical database, using 4-d optimal linear interpolation with anisotropic correlation scales related to topography (Loder et al. 1995). The spatial extent of various features is described, including low-salinity surface waters from the Gulf of St. Lawrence and coastal runoff, high-salinity Slope Water inflows in deep
channels, the seasonal upper-ocean temperature cycle, and increased summertime bottom temperatures in energetic tidal-mixing areas such as Georges Bank. Diagnostic model solutions indicate that the associated baroclinic circulation is the major contributor to the shelf-scale seasonal circulation in the Scotia-Maine region, including a seasonally-varying equatorward throughflow steered by the bank-basin topography (Hannah and Loder 1995). These fields are an important input to high-resolution model simulations in the Georges Bank region (Naimie et al. 1994; Naimie 1995) which include additional locally-important forcings such as tidal rectification. The fields are also a baseline for climatic variability evaluations.
As an initial evaluation of a decadal-scale climatic variation, 3-d seasonal-mean (composite) hydrographic fields are computed for groups of years representing the relatively-cool period in the
Scotia-Maine region during the late 1950s and 1960s, and the relatively-warm period during the 1970s and early 1980s. Average temperature and salinity differences between the two periods are
largest at depth along the shelf-break and in channels (e.g. Northeast) into shelf basins, and near the surface in the offshore Slope Water, with magnitudes about 5 degrees C and 1 unit respectively. Southwestward baroclinic flow along the shelf break is stronger during the cool period, particularly in winter, by about 2 Sv and 1 Sv on sections off Halifax and Georges Bank respectively. These results support the Petrie and Drinkwater (1993) suggestion that the anomalous conditions during the late 1950s and 1960s have a Slope Water origin related to increased strength of the Labrador Current. The present fields indicate that the anomalous conditions extend along the shelf break beyond Georges Bank, with widespread influences on the shelf hydrography, particularly in deep basins. Average temperature differences on southern Georges Bank are 1-4 degrees C, depending on season and location. Such significant decadal-scale changes during recent times with oceanographic and fisheries datasets provide the potential for evaluating the implications for the Georges Bank ecosystem of other possible climatic changes in the North Atlantic subpolar gyre.
References:
Greenberg, D.A., J.W. Loder, Y. Shen, D.R. Lynch and C.E. Naimie. 1995. Spatial and temporal structure of the barotropic response of the Scotian Shelf and Gulf of Maine to surface wind stress. J. Geophys. Res. (submitted).
Hannah, C.G, and J.W. Loder. 1995. Seasonal variation of the baroclinic circulation in the Scotia-Maine region. Proc. 7th Intern. Bienn. Confer. Phys. Estuar. Coastal Seas (submitted).
Loder, J.W., G. Han, C.G. Hannah, D.A. Greenberg and P.C. Smith. 1995. Hydrography and baroclinic circulation in the Scotian Shelf region: winter vs summer. Can. J. Fish. Aquat. Sci. (submitted).
Lynch, D.R., F.E. Werner, D.A. Greenberg and J.W. Loder. 1992. Diagnostic model for the baroclinic, wind-driven and tidal circulation in shallow seas. Cont. Shelf Res. 12: 37-64.
Naimie C.E. 1995. Georges Bank residual circulation during weak and strong stratification periods - prognostic numerical model results. J. Geophys. Res. (submitted).
Naimie, C.E., J.W. Loder and D.R. Lynch. 1994. Seasonal variation of the three-dimensional residual circulation on Georges Bank. J. Geophys. Res. 99: 15,967-15,989.
Petrie, B.D., and K. Drinkwater. 1993. Temperature and salinity variability on the Scotian Shelf and in the Gulf of Maine 1945-1990. J. Geophys. Res. 98: 20,079-20,089.
Monitoring the Inflows to the Gulf of Maine
Peter C. Smith
Bedford Institute of Oceanography, CA
Three physical oceanographic cruises (Oct. '93; June '94, '95) have been conducted in the eastern Gulf of Maine for the purpose of: 1) placing and recovering long-term moorings in Northeast Channel and off southwest Nova Scotia, 1) measuring the structure of inflow currents in Northeast Channel, and 3) surveying the hydrographic fields of the region. Basic processing of all current meter and CTD data has been completed, indicating a 90% data return from the moored data. Record mean currents from the moored instruments show persistent inflows of 5-10 cms-1 off Cape Sable and 5-15 cms-1 on the eastern side of Northeast Channel. Outflows of 10-20 cms-1 are found in the western side of the Channel, but these records are short. The expected annual cycles are also found in the inflows, featuring a fresh inflow maximum off Cape Sable in February and a pronounced, salty inflow in the Channel in August. However, strong interannual variability is also
noted in these signals, in part associated with the presence of warm core rings at the mouth of Northeast Channel. Furthermore, detided ADCP "snapshots" of the mean flow in the Channel indicate net transports consistent with previous estimates and maximum inflow and outflow currents of 40+ and 30+ cms-1 on the east and west, respectively.
Future plans include analyses of the wind-driven response, causes of interannual variability, the nature of "cross-over" events between Browns and Georges Banks, and seasonal variability in the baroclinic pressure field.
Evolution of Hydrographic Conditions on Georges Bank During the U.S. GLOBEC
1995 Field Season
David G. Mountain and Maureen H. Taylor
National Marine Fisheries Service, Woods Hole
Surveys of hydrographic conditions on Georges Bank were conducted each month from February through July 1995 as part of the broad scale studies component of the U.S GLOBEC Northwest Atlantic program. A grid of 38 stations was occupied using a CTD/flourometer instrument system.
The eastern portion of the Bank was directly influenced by waters from the Scotian Shelf from February through June. In addition during May Slope Water encroached onto the southern flank of the Bank to near the 80m isobath at the surface and the 70m isobath along the bottom. The Slope Water occupied the region of the Bank where the highest concentrations of cod and haddock larvae
are commonly found in May. The encroachment of Slope Water is believe to have been caused by a large warm core ring south of the Bank. By mid-June the Slope Water was gone and typical Georges Bank water covered the southern flank of the Bank.
Comparison with historic data indicates that the Georges Bank waters began the year somewhat warmer and saltier than normal. Through the spring the temperature and salinity anomalies progressively became cooler and fresher than normal, likely due to the presence of Scotian Shelf water.
Satellite and Drifter Evidence for Maine Coastal Current Flow Onto Georges Bank
Jim Bisagni
National Marine Fisheries Service, Narragansett
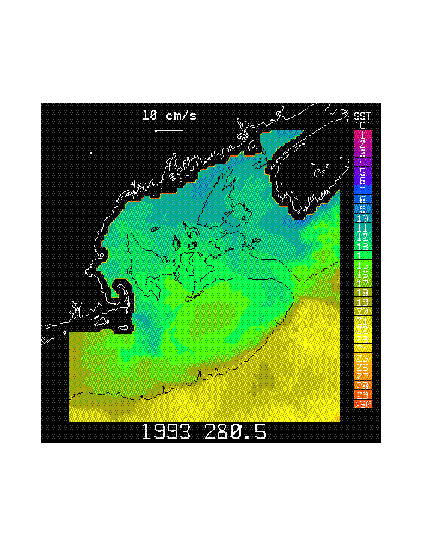
High-resolution, optimally-interpolated, satellite-derived sea surface temperature (SST) maps of the Gulf of Maine (GOM) and drogued Lagrangian drifters (drogue depths of 10 or 40 m) released in the northern GOM by N. Pettigrew (U. Maine) reveal the existence of the Maine Coastal Current (MCC) and show the MCC to be an extension of the colder and less-saline Scotian Shelf Water tongue which enters the eastern GOM south of Cape Sable, Nova Scotia. The MCC is manifested by decreased SST relative to waters outside the MCC during fall, spring and summer and mean along-coast (generally southwestward) current speeds of ~6-25 cm s-1 during spring-summer. SST contrasts across the MCC during winter are small due to deep convective cooling across the entire GOM. SST maps and drifter data reveal offshore extensions of the MCC located west of Jordan Basin, south of Penobscot Bay and south of Boothbay Harbor which may be related to offshore steering of the MCC by the Jordan Basin gyre and offshore spreading of the Penobscot and Kennebec/Androscoggin River freshwater plumes, respectively, in agreement with earlier observations and baroclinic numerical models. Data also show that MCC water may, at times, flow almost due south across the interior of the GOM and directly onto Georges Bank, to become entrained in the anticyclonic circulation on Georges Bank (Figure 1) and Figure 2).
Figure 1. Satellite-derived, optimally-interpolated sea surface temperature (SST) map from 8 October 1993 (yearday 280.5) showing a meandering southward extension of the cold Maine Coastal Current flowing onto Georges Bank. Ekman velocity at z = 10 m depth is close to zero. Also shown is a 10 cm/s reference velocity vector and a SST color bar.
Figure 2. Daily-averaged positions for drifter 22502 released within the Maine Coastal Current in northern Gulf of Maine on 29 April 1994 by N. Pettigrew, University of Maine, Orono, Maine. Drogue depth is 10 m. Also shown is the 200-m isobath (dashed). (Data courtesy of N. Pettigrew)
Prognostically Modeled Winter and Summer Circulation on Georges Bank
Christopher E. Naimie
Dartmouth College
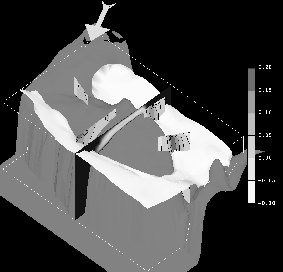
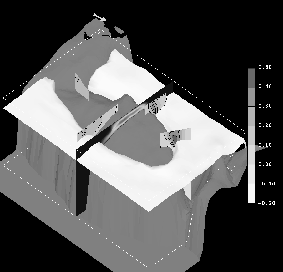
The seasonal extremes of the 3-D residual circulation on Georges Bank are studied by numerically computing the circulation during weakly (March-April, see Figure 1) and strongly (July-August, see Figure 2) stratified periods. The computations are performed using a prognostic (i.e. including tidal-time advection of the density field), free surface, nonlinear, finite element model on realistic Georges Bank topography. The dynamics considered result from the dominant M2 tide, baroclinic pressure gradients, wind stress, up-stream boundary conditions, and advanced turbulence closure. Comparison of these solutions with earlier diagnostic model results from Naimie et al. (1994) illustrates the importance of processes which could not be included in these earlier simulations (such as density evolution and advanced turbulence closure in tidal time, inclusion of nonlinear baroclinic effects, improved treatment of other nonlinearities, etc.).
During the weakly-stratified March-April period, the prognostic and diagnostic models predict similar global structure (with a recirculating transport on Georges Bank of 0.05 Sv), although there are local differences which result in the prognostic simulation agreeing more closely with the observed circulation (see Naimie (1995) for a more complete discussion).
During the strongly-stratified July-August period, the newly modeled processes intensify the residual circulation and result in recirculating transport on Georges Bank of 0.25 Sv, which is twice the diagnostic result. The level of agreement between modeled and observed residual circulation improves significantly for this period, with the prognostic model predicting around-bank currents as high as 46 cm s-1 in the frontal zone on the Northeast Peak. Analysis of the tidal-time density and current structure in the frontal zone on the Northeast Peak indicates important tidal frequency dynamics and features which are supported by observational evidence (see Naimie (1995) for a more complete discussion).
References
Naimie, C. E., J. W. Loder, and D. R. Lynch, Seasonal variation of the three-dimensional residual circulation on Georges Bank, J. Geophys. Res., 99(C8):15967--15989, 1994.
Naimie, C. E., Georges Bank residual circulation during weak and strong stratification periods - Prognostic numerical model results, J. Geophys. Res., in press, 1995.
Figure 1. Prognostically modeled winter (March-April) circulation on Georges Bank:
* The 26.25 Ut isopycnal intersects the topography on the southern flank of Georges Bank at roughly the 100 m isobath (white isosurface).
* The climatological wind stress of 0.0472 Pa is directed toward 121.4 degrees clockwise from true north (arrow above Nantucket Shoals indicates magnitude and direction).
* The vertical eddy viscosity varies from above 0.12 m2 sec-1 , at mid-depth on the crest of Georges Bank, to below .02 m2 sec-1, in the deeper surrounding regions (vertical cross-bank section).
* The strength of the around bank circulation peaks at 15-20 cm sec-1 for the northern flank, southern flank, and Northeast Peak (four vertical sections around the bank and legend (m sec-1)).
Figure 2. Prognostically modeled summer (July-August) circulation on Georges Bank:
* The 25.50 Ut isopycnal intersects the topography on the southern flank of Georges Bank at roughly the 80 m isobath (white isosurface).
* The climatological wind stress of 0.0138 Pa is directed toward 51.0 degrees clockwise from true north (arrow above Nantucket Shoals indicates magnitude and direction).
* The vertical eddy viscosity varies from above 0.12 m2 sec-1 , at mid-depth on the crest of Georges Bank, to below .02 m2 sec-1 , in the deeper surrounding regions (vertical cross-bank section).
* The strength of the around bank circulation varies significantly around the bank, reaching a maximum of 30-40, 20-30, and 40-50 cm sec-1 at the northern flank, southern flank, and Northeast Peak; respectively (four vertical sections around the bank and legend (m sec-1)).
Aspects of Shelfbreak Frontal Climatology
Chris Linder1, Glen Gawarkiewicz2 and
William Williams1
1MIT/WHOI
2 Woods Hole Oceanographic Institution
Background structure of the shelfbreak front on the south flank of Georges Bank will be described using a climatology developed from NODC data. Cross-shelf sections of temperature, salinity, and density were obtained using time averages of two months and by binning the stations by depth. Stations are averaged over roughly 100 km in the along-shelf direction, and in addition to Georges Bank fields for the shelf south of New England and off New Jersey were also computed. Because of the large gradients in bottom depth near the shelfbreak, relatively narrow spacing of the bins can be used in the frontal zone just offshore of the shelfbreak.
Despite the averaging over large space and time scales and hence various processes and frontal positions, the front shows fairly large near-surface velocities throughout the year, with velocity maxima ranging from 10 to 30 cm/s from the purely density-driven flow. The Georges Bank velocities in general are larger than those of the New England and New Jersey sections.
Mean positions of the front from the April/May time period were described and compared with a buoy trajectory provided by R. Limeburner and R. Beardsley. The buoy velocity was roughly 20 cm/s along the shelf edge as the buoy passed from the south flank of Georges Bank into the Middle Atlantic Bight.
Lagrangian Measurements of Near-surface Flow over Georges Bank
Richard Limeburner, Robert C. Beardsley, and Kenneth H. Brink
Woods Hole Oceanographic Institution
Almost every month during 1995, approximately five ARGOS-tracked surface drifters with holey-sock drogues centered at a depth of 10 to 15 m are being deployed over Georges Bank as part of the U.S. GLOBEC/Georges Bank program. The primary objective of this drifter program is to first characterize the near-surface Lagrangian flow over Georges Bank, especially its spatial and temporal structure and seasonal variability, and then integrate this physical information with other physical and biological measurements to help formulate a coherent description of the Georges Bank ecosystem.
Preliminary analysis plus previous drifter results from SCOPEX suggests that drifter behavior over the Bank is characterized by three regimes: wind-dominated, non-dispersive, and recirculation. In early February, 1995, strong northeast winds shifting to the northwest forced all (5) drifters over the Bank off-shelf where they became entrained in a warm-core ring. In March, strong northwest winds forced another drifter southwestward to the 100m isobath where it became entrained in the shelf/break frontal jet and moved quickly along the 100m isobath into the Middle Atlantic Bight . Many of the drifters deployed on Georges Bank between late February and June, 1995, remained within the region defined by the 60m isobath. The combined tracks of all these drifters generally describe the limits of the 60m isobath, except over the eastern side of the Great South Channel. Beginning in July, 1995, many drifters started to recirculate clockwise around the Bank along a narrow band generally located near the 60m isobath.
1Brookhaven National Laboratory
2Woods Hole OceanographicInstitution
There have been some 25 cruises on a variety of ships, on a variety of missions, transiting over and around Georges Bank during the GLOBEC Northwest Atlantic Program. On all the cruises ship-board acoustic Doppler current profilers (ADCP)s collected current velocity and acoustic backscatter intensity data. Both narrow (NB-) and broad-band (BB-) ADCPs were used in the study. Four ships have been used in the program, the Albatross, Endeavor, Seward Johnson, and Iselin. The ADCP types and arrangements differ on each of the ships; the Johnson had three different ADCPs all by itself. Some of the ADCPs were the older "narrow-band" variety, while some were the newer "broad-band" ADCPs. This diversity of equipment meant that we were unable to use the same data collection and processing methodology throughout as we had planned. For those ships equipped with the narrow-band ADCPs we were able to use, for the most part, RD Instruments' venerable ADCP DAS, version 2.48 (Endeavor and the last cruise on the Seward Johnson). On some of cruises with narrow-band ADCPs (Iselin and Seward Johnson) and for all those where the broad-band ADCPs were used (Iselin, Seward Johnson, and Albatross), data collection was controlled by RD Instrument's TRANSECT program.
Now that the flood of data is slacking off, the processing and analysis of this vast data set is underway. Of the 25 GLOBEC cruises of all sorts completed, the ADCP data has been processed for approximately 15 cruises. The processing is being done at both WHOI and BNL; all the TRANSECT data files are being processed at WHOI while all the PINGDATA files produced by the ADCP DAS are being processed at BNL. As the processing progresses, data products in the form of plots have been made available to GLOBEC investigators through the World Wide Web (http://www.oasdpo.bnl.gov/~oasdpo/mosaic/globec/).
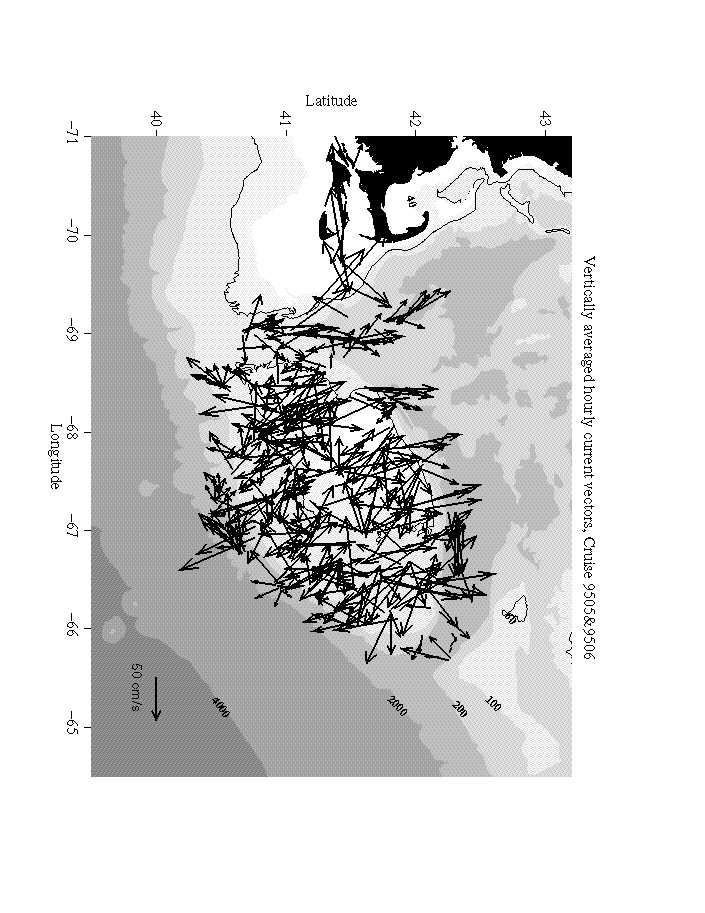
Some preliminary results from calculations of diurnal and semi-diurnal tidal components, the large-scale circulation over the Bank and the acoustic volume reverberation are presented here. Since Georges Bank currents are dominated by tides, one of the difficulties with the combined space/time character of ADCP data is to extract the tides from the mean and synoptic scale current fields. For this purpose we are using the detiding methods of Candela, Beardsley, and Limeburner (1992) which can decompose the observed currents into smoothly varying mean and tidal current fields. About three quarters of the current spatial and temporal variability from the ADCP observations over the Bank can be described using this type of model. The remaining variance represents smaller spatial and temporal variations due to topography, and seasonal and synoptic scale forcing for which the ADCP data will prove particularly useful.
Figure 1 shows the application of the methodology to two 1995 Albatross cruises, 9505 and 9506. As the figure shows, the background current field is completely overwhelmed by the tidal velocities. We are able to extract mean and semi-diurnal tidal current fields which at least qualitatively conform to the existing Eulerian estimates. The semi-diurnal tidal magnitudes and orientations appear to be closer to the observations in than the mean field which seems too small. We will be combining the from all the cruises as well as moored instrumentation to construct a much more accurate semi-diurnal and diurnal tidal description for detiding the ADCP currents.
One of the more useful measurements made possible by the ADCP is an estimate of the acoustic volume reverberation all along the cruise track. It has been shown in a number of studies that the volume reverberation from a single frequency instrument like the ADCP can be used to estimate the zooplankton biomass. These estimates are not as accurate as those obtained with higher frequency and multi frequency acoustic devices, optics or nets, but these limitations are compensated for by greater range and ease of use. We have continued to refine the processing techniques and the need to use the backscatter intensity data over the large shallow areas of the Bank has led to a re-examination of processing methods originally developed for deep water applications. A provisional method relaxes the percent good criterion progressively toward the surface above 50 meters while tightening the percent good criterion below that depth, from our usual 92% to 95%. Because of potential miss-alignments of the ADCP AGC amplifier, the top bin is also being rejected.
The Application of Oxygen Isotope Analysis for Water Source Identification
Robert. W. Houghton and R. G. Fairbanks
Lamont Doherty Earth Observatory
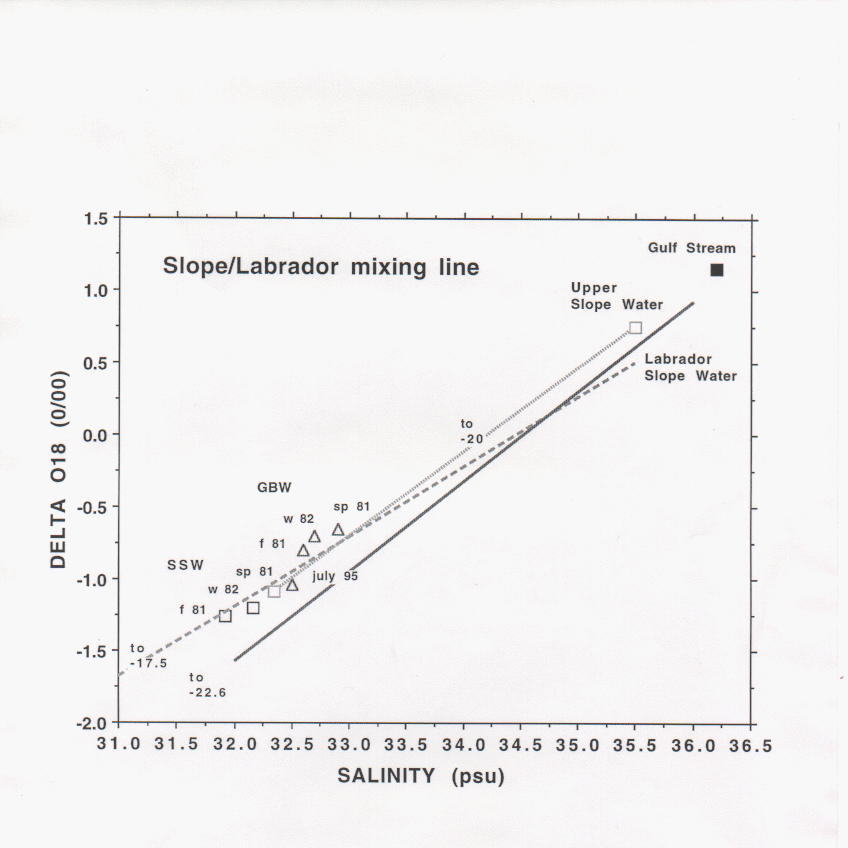
The isotopic ratio, 18O/16O, is a conservative tracer useful for distinguishing freshwater sources especially along the east coast of North America where the value of this ratio for meteoric water diminishes with latitude so that river watersheds can be identified by their isotopic ratio. It is found that the isotope and salinity values of Slope Water along the NE continental margin define a binary mixing line between Gulf Stream water and Labrador meteoric water (delta 18O = -22 0/00, Figure 1). Deviations from this regression have been used to identify local freshwater sources (Fairbanks, 1982). In the Middle Atlantic Bight (MAB) the regression curve between upper Slope Water (USW) and Scotian Shelf Water (SSW) projects to freshwater at delta 18O = -20 0/00 suggesting that the Labrador Sea margin is the primary freshwater source throughout the NE continental margin (Chapman et al., 1986; Chapman and Beardsley, 1989).
On the Scotian Shelf, Georges Bank, and in the Gulf of Maine isotope-salinity values fall along a mixing line between Labrador Slope Water (LSW) and freshwater at delta 18O = -17 0/00. Although values for Georges Bank Water (GBW) at the Bank crest appear to fall along this mixing line analysis of Gulf of Maine data (Fairbanks, 1983) shows that GBW predominantly has the water properties of Wilkinson Basin and is the result of complex mixing processes of SSW, LSW, and local river sources within the Gulf of Maine. These appear as distinct mixing lines in isotope-salinity plots in contrast to the more confused patterns in equivalent temperature-salinity plots. The isotope-salinity value of GBW undergoes seasonal and interannual variations. From Al9508, we find that GBW in July 1995 was fresher than the MARMAP 1981 and 1982 observations. An objective of planned research is to determine if these changes are due solely to changes in the SSW incident directly onto the bank or to changes in the circulation and mixing processes within the Gulf of Maine.
The apparent absence of a distinct signature of St. Lawrence River water seems curious since estimates of freshwater flux (Loder et al., 1995) suggest that the St. Lawrence River contributes 1/3 to 1/4 of the freshwater entering the Scotian Shelf. It can be shown that mixing within the Gulf of St. Lawrence of this fraction of freshwater with water on the Labrador-Gulf Stream mixing curve would produce a water type that appears to have a freshwater source with delta 18O = -17 0/00. This becomes the end member for the mixing line on the Scotian Shelf. Thus the identification of Labrador as the sole freshwater source on the outer shelf in the MAB is misleading.
On the NE continental margin the oxygen isotope is a powerful tracer for distinguishing freshwater sources. It can distinguish SSW directly incident onto Georges Bank from freshwater derived from the Gulf of Maine. It can also distinguish saltier USW, with a Gulf Stream-Sargasso Sea origin, from
LSW.
References:
Chapman, D.C. and R.C. Beardsley, 1989. On the origin of shelf water in the Middle Atlantic Bight. J. Phys. Oceanogr., 19, 389-391.
Chapman, D.C., J.A. Barth, R.C. Beardsley, and R.G. Fairbanks, 1986. On the continuity of mean flow between the Scotian Shelf and the Middle Atlantic Bight. J. Phys. Oceanogr., 16, 758-772.
Fairbanks, R.G., 1982. The origin of continental shelf and Slope Water in the New York Bight and Gulf of Maine: evidence from H218O/H216O ratio measurements. J. Geophys. Res., 87, 5796-5808.
Fairbanks, R.G., 1983. Isotopic origins of Georges Bank water. Special document for the National Marine Fisheries Services (NOAA), Woods Hole, MA.
Loder, J.W., B. Petrie and G. Gawarkiewicz, 1995: The coastal ocean off Northeastern North America: A large-scale view. (submitted for publication in THE SEA)
Figure 1: Representative binary mixing curves, with the zero salinity intercept indicated, for the Scotian Shelf to the Middle Atlantic Bight region from Fairbanks, 1982 (solid line); Chapman et al., 1986 (dotted line); Fairbanks, 1983 (dashed line). Depth averaged isotope-salinity values for particular years and seasons are shown for SSW (squares) and GBW (triangles).
Three-Dimensional Distribution and Abundance of Zooplankton on Georges Bank, 1939-41, with Comments on the Effects of Storm Events
Steve Bollens, Mary Landsteiner, Cabell Davis, and Andy Solow
Woods Hole Oceanographic Institution
There are three overall objectives to this research project: 1) analyses of a newly uncovered and largely unpublished data set from the RV Atlantis (1939-1941) for distribution, abundance, and life histories of the dominant zooplankton on Georges Bank (it is worth noting that these are the only 3-D, bank-wide extant zooplankton data from Georges Bank); 2) investigation of storm effects on the distribution and abundance of zooplankton; and 3) comparison of Atlantis data with more recent data sets (e.g., MARMAP) to investigate possible longer-term (i.e., inter-decadal) changes in zooplankton abundance. With regard to the first two objectives, the following highlights are meant to summarize our research findings to date; results relating to the third objective are summarized in the abstract by Landsteiner et al. under the "hot topics" session.
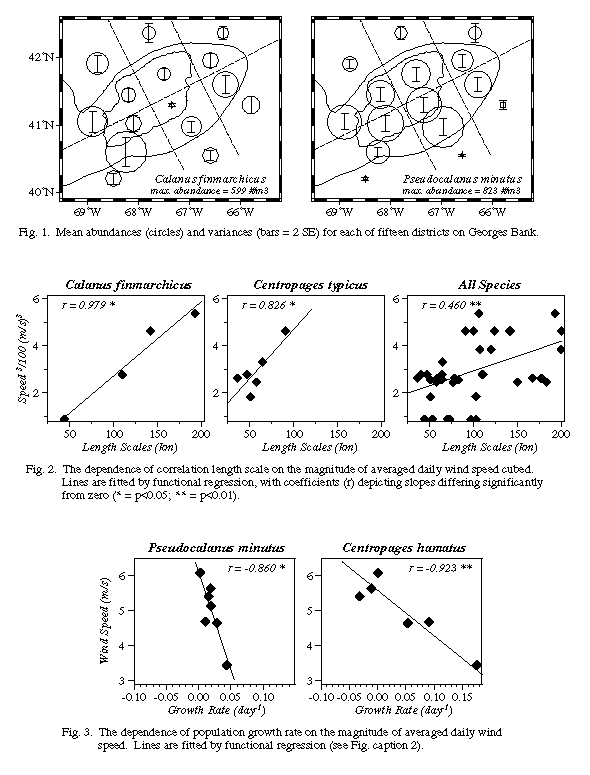
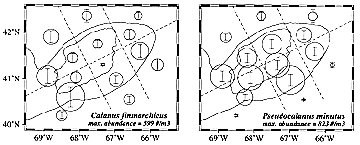
* Bank-wide horizontal distributions varied considerably between seasons, years and region, and often did so in a species-specific manner (e.g., Figure 1). For instance, regions of higher than average abundance for the dominant species were as follows: Sagitta and Centropages hamatus <60 m; Calanus and C. typicus between 60-100m; Pseudocalanus <100m; Metridia > 100m. These patterns were similar to those described by other investigators using more recent (e.g., MARMAP) data.
* Vertical distributions of zooplankton on the bank during 1939-41 also varied enormously - between species, between developmental stages, between cruises, and between regions of the bank - with changes in DVM behavior (e.g., reverse vs. normal) occurring in several species. For instance, daytime depth of Calanus increased with developmental stage such that early copepodid stages were typically non-migratory in the surface but later stages were undertaking normal DVM, although a statistically significant reverse migration in adult Calanus was observed on at least one date. For Pseudocalanus, changes in DVM behavior occurred between seasons (e.g., during spring 1940, reverse DVM in March-April, non-migratory in April-May, and normal DVM in June) and between regions (reverse DVM in waters < 100 m and normal DVM in waters >100m during May-June 1941).
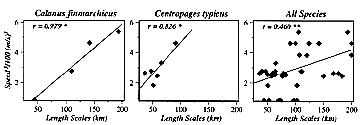
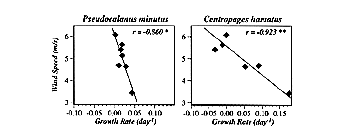
* Horizontal patchiness (e.g., correlation length scales [CLSs]) of zooplankton on Georges Bank appear to be affected by storms (wind events). For instance, daily wind speed cubed, a common proxy for mixing intensity, averaged over 30 days prior to each cruise, was positively significantly correlated with the CLSs of Calanus, Centropages, and all species combined (Figure 2). The five other abundant taxa considered all showed positive but insignificant correlations. Thus storminess would appear to break down (homogenize) small-scale patchiness and thereby increase the length scales at which zooplankton abundances are correlated.
* Population growth rates of zooplankton on Georges Bank also appear to be affected by storms (wind events), with those species dominant in the shoal regions of the bank being most strongly influenced. For instance, realized rate of population increase between cruises was significantly negatively correlated with the magnitude of average daily wind speed calculated over the same periods for PseudoCalanus and C. hamatus (Figure 3), both of which are shoally distributed on or near the peak of the bank; species more deeply distributed off-bank (e.g., Calanus and Metridia) showed no such dependence. However, specific events might be expected to effect all species - a comparison of the May-June inter-cruise period for 1940 and 1941 shows that in the high wind year (1940; mean = 5.6 m s-1) population growth rates for the 10 dominant taxa were in all cases lower than in the low wind year (1941; mean = 3.4 m s-1). Thus storminess would appear to negatively affect zooplankton abundances on Georges Bank, perhaps by advecting animals off the bank.
Figure 1. Means (circles) and variances (vertical bars = 2 SE) of the abundance of Calanus and Pseudocalanus for each of fifteen districts on Georges Bank.
Figure 2. The dependence of correlation length scale (km) on the magnitude of averaged daily wind speed cubed. Lines are fitted by functional regression, with coefficients (r) depicting slopes differing significantly from zero (* = p<0.05; ** = p<0.01).
Figure 3. The dependence of population growth rate on the magnitude of averaged daily wind speed. Lines are fitted by functional regression, with coefficients (r) depicting slopes differing significantly from zero (* = p<0.05; ** = p<0.01).
Spatial and seasonal patterns in abundance and age-composition of Calanus finmarchicus and Pseudocalanus spp. in the Gulf of Maine and on Georges Bank: 1977-1987
Carol J. Meise1, Wallace Morse1, John Green1 and Edward Durbin2
1National Marine Fisheries Service, Narragansett
2University of Rhode Island
Spatial and seasonal patterns in abundance and age-composition of Calanus finmarchicus and Pseudocalanus spp. on the northeast continental shelf, based on 333 mesh MARMAP surveys (1977-1987) were described. A large part of the spatial and seasonal variation in C. finmarchicus abundance and age structure appears to be tightly coupled to major hydrographic regimes and to major circulation patterns in the region. There was a sharp ecotone between well-mixed Georges Bank and the Gulf of Maine as defined by C. finmarchicus abundance patterns and life history distributions. The ecotone is present year-round, but is most apparent during the stratified season (May-October), when thermohaline density gradients and the near-surface current jet along the northern flank are generally strongest. The Gulf of Maine had the highest abundances of C. finmarchicus, and lowest spatial and seasonal variation in the region while tidally mixed Georges Banks displayed the opposite pattern. This indication of stable population centers in the Gulf of Maine would make it a major source of Calanus in the region, particularly during March-April. Distributional patterns also suggest a strong Calanus influence from Scotian Shelf water in northern Gulf of Maine and on the southern flank of Georges Bank. Calanus distributional patterns do not display a sharp ecotone between well mixed Georges Bank and the GOM. In-shore GOM has the highest abundances of Pseudocalanus, while lower abundances are found in deeper GOM waters. Pseudocalanus abundance on Georges Bank is lowest from Sept-Dec and peaks in May-June.
Seasonal abundance of copepodites stages 1-5 for Pseudocalanus and 1-3 for C. finmarchicus from 165-053 um mesh are described for the Georges Bank-Gulf of Maine region. These data show a continual (throughout the year) presence of younger stage copepodites.
Spatial and Temporal Patterns in Abundance of Invertebrate Predators from MARMAP Data, 1977-1987
Barbara Sullivan1 and Carol Meise2
1University of Rhode Island
2National Marine Fisheries Service, Narragansett
Chaetognaths, primarily Sagitta elegans, were the most abundant and widespread predator on Georges Bank during 1977-1987 MARMAP surveys. They were present in 79% of samples collected and on average were an order of magnitude more abundant than other predators. Cnidaria
(probably hydrozoan and scyphozoan medusa), euphausiid shrimp and gammarid amphipods were the next most abundant groups. Ctenophores and adults of large, rapidly swimming crustacean predators were probably underrepresented in MARMAP samples. Diversity and abundance of predators was highest in the shoal well-mixed region of the bank. In this region predator abundance was positively correlated with abundance of Calanus finmarchicus but was inversely correlated with Calanus abundance in deeper waters.
Chaetognaths and cnidaria were more abundant in 1978-1979 than in other years; there was a statistically significant decline in abundance of chaetognaths over the 10 year period. Abundance of chaetognaths in summer was inversely correlated with average temperature during the previous winter; abundance of cnidaria in summer decreased with increasing summer temperatures. (See Figure 1, following page, and Figure 2, below.)
Recruitment and Growth of Calanus finmarchicus and Pseudocalanus spp on Georges Bank during late spring
Edward Durbin1, Jeff Runge2, Robert Campbell1, Peter Garrahan1, M. Casas1, S. Plourde2
1University of Rhode Island
2Institut Maurice-Lamontagne, CA
Population dynamics of the dominant copepods on Georges Bank were investigated during late May-early June 1994 as part of the US-GLOBEC Georges Bank Program. ARGOS/GPS drifters were deployed on the northeast peak, in the shallow well-mixed region, and on the southern flank of Georges Bank. These were used to track zooplankton populations for up to three days at each site. MOCNESS and plankton pump sampling provided a complete description of the age structure of the dominant copepods at each site. Differences in egg production rates and age structure of Calanus finmarchicus between the northeast peak and the southern flank, coupled with the drifter tracks, suggest a pattern of advection of copepods onto the bank, increased egg laying as they encounter higher food levels, and then the subsequent advection of this developing cohort along the southern flank. Stage durations measured in mesocosms on board ship varied with stage at a given site. Nauplii and young copepodites generally developed quite rapidly, and in some cases near estimated maximum rates, at all sites. In contrast, late stage copepodites of Calanus and Pseudocalanus were developing very slowly. Results indicate that environmental conditions for growth and production on Georges Bank vary significantly for different developmental stages of the dominant copepods.
GLOBEC Broad-scale larval fish feeding studies on Georges Bank: a preliminary analysis
Jack Green, Toni Chute and Rebecca Jones
National Marine Fisheries Service, Narragansett
During April and May 1995 an array of 24 stations was sampled after the wide area coverage had been completed under the GLOBEC Georges Bank Broadscale sampling plan. These stations, arrayed in a 'starburst' pattern approximately 30 nm in diameter, provided samples for analyses related to the growth and survival of larval cod and haddock. Twelve 1m2 MOCNESS samples were analyzed for larval cod and haddock gut contents. Samples (150 m mesh) were sorted for larvae and the guts examined for identification and enumeration of prey items along with a variety of larval morphological measurements to characterize changes in prey preference with larval development.
The April 'starburst' was located well to the west of the highest larval fish distribution as determined by the Bank-wide survey but coincided with the location of the center of larval fish distribution located in the May cruise; in effect providing a resampling of the same location one month later. In April the abundance of cod and haddock larvae in the samples was much higher than in May. The size ranges of cod and haddock in April are similar, with most of the fish ranging between 3 and 10mm SL and a peak for both species between 6 and 7 mm SL. By May the abundance was much lower and the size range increased to 4-23mm for cod and 4-10mm for haddock larvae. Length frequencies observed for both cruises are shown in Figure 1.
Although 90 percent of the larvae examined contained prey in the guts, there was no correlation between the diel distribution of larvae and occurrence of food in the guts. Gut contents were identified to the lowest possible taxon and showed a wide range of prey (Figure 2). In general, it appears that both species were eating the same sorts of prey with naupliar stages of calanoids and Oithona as well as invertebrate eggs comprising the most abundant categories in April. By May, the larger size of the larvae appears to allow predation on larger organisms as is shown by the increased occurrence of copepodites in the gut. However there is still significant predation on smaller groups indicating that the range in size of the prey has increased rather than shifting from smaller to larger prey. Veligers, which are likely to be indigestible, Coscinodiscus, and sand grains were found in a few of the stomachs.
The complete study of the stomach contents of larval cod and haddock, will include analysis of the entire sample array for all 1995 Broadscale cruises and will include analysis of prey availability and selectivity as a function of larval development and environmental conditions.
Figure 1:
Figure 2:
Biological and physical evidence of source regions and transport patterns of zooplankton on Georges Bank
Ann Bucklin, C.C. Caudill, and S.B. Smolenack
University of New Hampshire
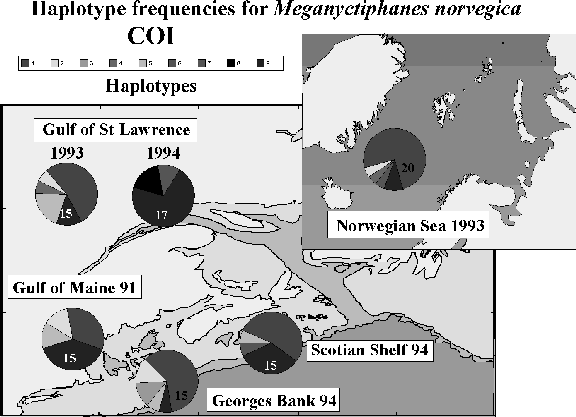
Analysis of the base sequence variation of mitochondrial DNA (mtDNA) for the copepod, Calanusfinmarchicus, and the euphausiid, Meganyctiphanes norvegica, revealed differences in the levels of variation among three gene portions, based on the numbers of haplotypes and the genetic distance between haplotypes. 16S rRNA was most conserved, cytochrome oxidase I (COI) was somewhat more variable, and cytochrome b (CYB) was most variable. Calanus finmarchicus in the NW Atlantic is a single, genetically homogeneous population based on 16S rRNA (Bucklin and Kocher, 1995). The validity of mtDNA as a marker of population structure was borne out by parallel analysis of the copepod, Acartia tonsa, found in estuaries along the east coast of the U.S. This species comprised three deeply divergent groups of lineages, exhibited higher nucleotide diversity than C. finmarchicus or M. norvegica, and showed significant differences in haplotype frequencies between adjacent estuaries. Meganyctiphanes norvegica revealed significant differences in COI haplotype frequencies in the NW Atlantic
(Figure 1). CYB haplotypes were signficiantly divergent on either side of the N. Atlantic based on an analysis of molecular variation (AMOVA). MtDNA genes differ in patterns of evolution in ways that affect their usefulness as markers of ecological processes in marine populations. Genes of moderate variability may prove most useful for population genetic studies, since using a highly variable marker results in many unique individuals that are not geographically
informative.
Thus, C. finmarchicus constitutes a single population (genetically defined) from the Gulf of St. Lawrence to the Gulf of Maine. Within this homogeneous domain, dispsersal pathways will be traced by patterns of genetic similarity among the sampled individuals and knowledge of circulation patterns. This approach may help test our hypothesis that C. finmarchicus of the Gulf of St. Lawrence is a primary source of recruitment for the species in the NW Atlantic.
References
Bucklin, A. and T.D. Kocher. 1995. Source regions for recruitment of Calanus finmarchicus to Georges Bank: evidence from molecular population genetic analysis of mtDNA. Deep-Sea Res. (in press)
Figure 1. Haplotype frequencies for the euphausiid, Meganyctiphanes norvegica, in samples from Norway, Gulf of St. Lawrence, Scotian Shelf, and the Gulf of Maine from 1991 to 1994. A 400 base-pair region of cytochrome oxidase I (COI) was sequenced for 97 individuals. [Sample sizes are indicated by numbers on the bottom of haplotype frequency diagrams. Haplotype "P" is the pooled frequency of all unique haplotypes.]
Relationships Among Springtime Hydrography, Phytoplankton Chlorophyll, Nutrients and Cod Larvae on Georges Bank in April and May of 1993 and 1994
David W. Townsend
University of Maine
Shipboard surveys of larval cod and hydrography, including measurements of phytoplankton chlorophyll a and nitrite+nitrate, were conducted on the R/V Columbus Iselin on 20-29 April and 17-26 May of 1993, and 7-15 April and 12-20 May of 1994. The temperature and salinity fields in 1993 were influenced along the southern edge of the Bank by the close proximity of warm, salty Gulf Stream ring (or Slope) water, and on the Northeast Peak by an intrusion of relatively fresh, cold, low-nutrient and low-chlorophyll surface waters of Scotian Shelf origin. The Scotian Shelf Water (SSW) could be tracked between the April and May 1993 surveys, as it advected clockwise around the perimeter of the Bank. The larvae were concentrated in two patches, which were also advected around the Bank (patterns of larval drift are discussed in Townsend and Pettigrew in the Deep Sea Research volume). Surface nutrients in May 1993 (>2.0 µM-N; not measured in April) extended as a broad band running north-south along the Northeast Peak. Chlorophyll a was highest on the Northern edge and Northeast Peak, in both April (>10 µg L-1) and May (>6 µg L
1) of 1993; larval cod densities appeared to co-vary with the chlorophyll distributions in both months, in that larvae were more abundant in the low-chlorophyll waters, perhaps associated with the edges of SSW. Larval densities were reduced by about 75% between surveys in 1993, and by more than 90% in 1994. Intrusions of SSW was also observed in April of 1994, but it was less dramatic than the previous year. There was again Gulf Stream ring/Slope Water on the southeastern edge of the Bank. Nutrients in April 1994 were concentrated along a band (4-8 µM
N) that ran seaward of the 60m isobath, from the Northern Edge to the Southern Flank. In May nutrients were similarly distributed, but they were reduced to about two thirds their concentrations in April. Chlorophyll was highest over the shallower portions of the Bank in both April (6-20 µg L-1) and May (1-4 µg L-1) of 1994. Cod larvae in April 1994 were most abundant on the southeastern flank of the Bank, where they formed a single broad patch. In May the larvae were much less abundant and more diffuse about the Bank; they were not concentrated in coherent patches, and their transport between surveys could not be followed.
These cruise results demonstrate that winter concentrations of nutrients on the top of the Bank (inside the 60m isobath) become depleted by the spring phytoplankton bloom, which likely begins in the late winter (March-April) and does not depend upon vertical water column stability (sensu Townsend et al., 1994; Deep Sea Res. I, 41: 747-765). Following the bloom (late April - May), chlorophyll distributions are limited to the northern edge and frontal periphery of the Bank, between the 60 and 100m isobaths; those phytoplankton populations are most concentrated on the Northeast Peak, and appear to be supported by cross frontal fluxes of new nutrients that upwell onto the northern edge and Northeast Peak, and then flow around the Bank toward the Southern Flank. Although patches of relatively high chlorophyll concentrations are also found in May on the Southern Flank and on the top of the Bank, they appear to be supported by regenerated nutrients. Cross-Bank vertical sections on the Southern Flank suggest that off-Bank sources of "new nitrogen" (NO3) there are quickly utilized as those waters are entrained and upwelled onto the relatively broad southern flanks of the Bank, and those fluxes do not reach the central portions of the Bank.
These results suggest that primary and secondary production on Georges Bank is nitrogen-limited, and that this limitation can be characterized by region on the Bank. As has been suggested earlier by Horne et al. (1989; Scient. Mar. 53: 145-158), "new primary production" [that dependent on NO3] is more important on the northern half of the Bank and in the frontal regions; overall primary production is greatest there as well. Primary production on the top of the Bank and on the southern half, on the other hand, is more dependent upon "regenerated production" [that utilizing NH4]. For this reason, it follows that secondary production on Georges Bank is also nitrogen
limited, and that zooplankton production (addition of new biomass) is higher on -- and perhaps limited to -- the Northern Flank and Northeast Peak, where new nitrogen [NO3] is supplied. Zooplankton biomass, on the other hand, is likely greater downstream in the "older" waters on the Southern Flank and on the top of the Bank, where primary production is lower, and nitrogen
limited. The higher standing stocks of zooplankton at these locations are likely limited to maintenance metabolism only, which is fueled by regenerated primary production; these populations are unlikely to produce new protein. It then follows that successful cod spawning on Georges Bank depends upon larvae being transported to such regions that are enriched with high standing stocks of their zooplankton prey items, which are in turn produced some distance upstream.
Bank-wide Distribution, Sources and Suspension of Planktonic Hydroids, 1994 and 1995
Barbara Sullivan1, Steve Bollens2, Stephanie Concelman2, Erich Horgan2, Mari Butler2, Larry Madin2, Donna Van Keuren1
1University of Rhode Island
2Woods Hole Oceanographic Institution
As a result of the 1994 field season, planktonic hydroids, primarily of the species Clytia gracilis, were newly recognized as an important component of Georges Banks zooplankton community. Laboratory experiments and gut content analyses indicated that they could consume 50-100% of the daily production of copepods and may also directly prey upon cod larvae.
During 1995 intensive studies of both planktonic and benthic distribution of hydroids were designed to determine how hydroids accumulate and are maintained in the plankton. Planktonic hydroids were
broadly distributed over the southern flank of the Bank with highest abundance over the central shoal region, especially where the shelf/Slope Water front intersects the bottom. Benthic colonies exhibited a very different distribution being most abundant on the North East Peak. These observations are consistent with the idea of there being an upstream source of benthic hydroids that interacts with the clockwise gyre circulation on the Bank to seed and maintain a large population of planktonic hydroids in the central, shoal region. Additionally, laboratory investigations on growth and life history characteristics of planktonic hydroids indicate an important role for mixing in addition to food availability. Clytia gracilis seems peculiarly adapted to thrive in the plankton of Georges Bank. (See
Our basic understanding of the usual seasonal pattern of life history events in Calanus finmarchicus was provided in a 1934 paper by S. Marshall, A. G. Nicholls and A. P. Orr (1934, JMBAUK, 19: 793-828). Their study considered the population in the Clyde Sea area of western Scotland. Half of the year is spent by the Calanus stock resting at depth in the fifth copepodite (C5) stage. In February this resting stock completes its maturation as generation 'Go', C6 males appearing before females. The females ascend to the upper water column and commence spawning of generation 'G1' when the spring bloom provides copious food to support egg production. G1 requires about 2 months to develop, maturing immediately without rest and producing G2. G2 develops slightly faster, then in July or early August returns as C5 to depth to repeat the cycle.
Variants on this basic theme have been noted, many of them reviewed in Marshall and Orr's book (1955, The Biology of a Marine Copepod, Calanus finmarchicus (Gunnerus), Oliver and Boyd, Edinburgh, 180 pp.). For example, fjord populations north of central Norway have strictly one generation per year, C5 of G1 returning to depth. Kaartvedt (pers. comm.) suggests this is because continuous summer illumination provides no time near the surface when visual predators cannot see Calanus. Resting stocks at most locations turn out not to be entirely C5, but to include a fraction of C4. The depths at which C5 rest vary greatly across the range. In New England Slope Water the resting stock centers around 470 m. In the basins of the Gulf of Maine and Scotian shelf it resides just above the basin bottom. In the eastern Atlantic it is at and below 800 m.
The Georges Bank program has observed a number of new variants on the standard theme. In November, 1994, Durbin, Miller et al. observed a sizeable fraction of the C5 stock in the upper 30 meters of the water column, actively feeding both day and night. These animals showed no preliminary signs of breaking their rest, such as gonadal or adult tooth development. Probably they found a period in which the surface was not patrolled by visual predators and took advantage of it. Meise has determined from MARMAP samples that a small fraction of the Gulf of Maine stock continues active reproduction and development right through the year. Apparently the resting stage is not obligatory. We have observed in both 1994 and 1995 seasons that some fraction of each spring generation (G1, G2, possibly G3) initiates rest, while another group matures and spawns immediately. We don't know what determines which of these paths a given individual will take. Genetically determined behavior based on timing cues for the season (e.g., photoperiod changes) could vary among individuals, or they could somehow receive different cues from the habitat. We do not, yet, have a method for testing these hypotheses.
Major life history issues currently under investigation by us and by European participants in the TransAtlantic Studies of Calanus program include evaluations of (1) the extent to which developing stocks over shoal continental shelves (Georges Bank, the North Sea) are supplied from deep basins or offshore and (2) the extent to which the highly productive shelf populations of spring return resting C5 to deep waters. (3) It is likely that some genetically male Calanus mature as females. Georges Bank and Norwegian fjord stocks are being examined to determine frequency and seasonal dependence of this sex change. (4) Studies throughout the range will determine the dependence of fecundity in Calanus upon seasonal timing, food availability, temperature, and other factors. We are fine tuning our paradigm for the life history of Calanus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![Figure 4. Calanus finmarchicus, C5. Video estimates of oil sac volume vs. prosome length for all specimens recorded in June. Other months showed a similar pattern. Dashed line is an approximate upper limit on oil sac size as a function of body length. Seasonal Patterns In Enzyme Activities and Condition of Larval Fish and Copepods on Georges Bank M. Elizabeth Clarke University of Miami The focus of out research was to look at a variety of species in a variety of locations to test the utility of glycolytic enzyme assays as a method for rapidly assessing the condition of zooplankton and ichthyoplankton. We have collected samples in a variety of habitats and on many species to this end. As part of this wider study designed to develop a method of assessing condition of ichthyoplankton and zooplankton, we analyzed samples collected during the Georges Bank program. We believe that activities of enzymes of metabolic pathways that generate energy (e.g., lactate dehydrogenase (LDH) of anaerobic glycolysis and citrate synthase (CS) of the Krebs cycle) can be practical indicators of the nutritional status and growth, respectively, of larval fish and other zooplankton. First, these techniques are easy to apply, requiring only a sonicator, microcentrifuge and recording spectrophotometer as specialized equipment. Second, enzyme activities are exquisitely sensitive to feeding regime in adult fish and enzyme activities scale with body length in adult fish and larval in a manner that reflects thrust requirements for locomotion. Additionally, that activities correlate well with oxygen consumption has suggested that the difference between LDH and CS in larval fish and zooplankton represents the scope for activity (i.e., the fish's potential to increase its activity over basal levels to its maximum). By applying these enzymatic assays in both laboratory experiments and in the field we find that the factors which affect activity are food abundance, sex, age or stage. (Figure 1) We have concluded from this that enzymatic activities are practical proxies for condition in larval fish and zooplankton. Using enzyme activities, condition can be resolved at the level of the individual organism rather than from a multi-species homogenate. While we have sampled animals in South Florida, the South Atlantic Bight, Gulf of Mexico and the South Atlantic Bight as well as on Georges Bank. For Georges Bank, we have the widest seasonal coverage. We have participated in cruises at two time periods in both 1993, 1994 and one time period in 1995. We have conducted experimental studies on larval cod to illustrate the sensitivity of the technique on larval fish. CS, LDH and NDPK activities all are significantly different between fish fed 50 food items per liter vs those fed 200 food items per liter for 10 days. In collaboration with Elaine Calderone at NMFS, Narragansett, we tested the enzymatic activities of cod larvae 14 days old and starved for 3 days. There are significant differences in LDH and NDPK after starvation for 3 days. Field studies of enzymatic activity indicate that there is some variability in the condition of larvae and copepods which are probably related to smaller scale features in the local oceanography. Seasonal patterns in condition are evident. When bank-wide surveys were analyzed in cod less than 6 mm collected in 1993 an 1994, the CS and LDH values April and May are significantly different in both years with April higher. There is also a difference in condition of cod between 1993 and 1994. For the same time period, condition is lower in 1994 than 1993. For Calanus (CV) we see a different pattern. April condition is higher thatn May in 1993 and in 1994 condition is at the same high level as was seen in April 1993 in both April and May 1994. There are also differnces in condition of larval cod collected in an "on-bank" vs. "off-bank series" of 1 m2 Mocness tows in 1993 and 1994 (Figure 2). As was evident in the bank-wide sampling the LDH activities are higher in 1993 than 1994. Further, the "off-bank" samples are higher in condition than the "on-bank" samples in the same year. Figure 1: Figure 2: Acoustical estimation of the distribution of zooplankton biomass on Georges Bank Peter Wiebe1, Mark Benfield1, Tim Stanton1, Cabell Davis1, and Charles Greene2 1Woods Hole Oceanographic Institution 2Cornell University High frequency (420 kHz) sound data in combination with Video Plankton Recorder (VPR)and MOCNESS zooplankton data were collected in the physically stratified area on the southern flank of the Georges Bank in May 1992. The echo sounder was mounted in a towed V-fin with its transducers downlooking and was towed from the R/V Albatross IV along a trackline that defined a grid approximately 2 by 4 kilometers. Volume backscattering data were collected from near surface to the bottom (~ 80 m). On the last segment of the trackline, a 1- m2 MOCNESS was towed obliquely and provided samples in 10 meter intervals from 50 m to the surface. The VPR was towyoed (0 to 70 m) from a second ship, R/V Endeavor, which moved along similar trackline displaced by about 180 m. The MOCNESS and VPR provided quantitatively comparable estimates of the abundance and size of the most abundant taxa (copepods, and pteropods) present in the grid. Using the assumption that the relative proportions of taxa remained fixed at constant depth, volume backscattering was used to scale the numbers of individuals present in the ensonified volume. These values were then partitioned into numbers per taxa based on the relative proportions of individuals observed in the VPR and MOCNESS samples. Estimates of wet weight were computed from the numbers per taxa and knowledge of their size in the grid area. The pattern of estimated biomass distribution differed in significant ways from the pattern of acoustical volume backscattering. Furthermore, using current theoretical models for volume backscattering, estimates of the contribution of each taxa to the 420 kHz volume backscattering were computed based on their numbers and size as a check to see how well the acoustic predictions matched the acoustic observations. The techniques being developed to use high frequency acoustical backscattering data to estimate zooplankton biomass will be applied to the broad-scale acoustical data collected in June 1994, February 1995 and July 1995. Climatological Box Modeling of the Gulf of Maine and Georges Bank Region Wendell S. Brown and Frank L. Bub University of New Hampshire Climatological Hydrography The AFAP hydrographic data have been divided into four density layers which approximately correspond to the principal water masses in the Gulf of Maine (surface, intermediate, bottom and Atlantic Slope Water). The monthly climatological data for the period 1970-89 were interpolated onto the 7 km Greenberg (1983) model grid of the region using Objective Analysis (OA) techniques (Carter and Robinson, 1987). The results clearly document the annual evolution of each layer's thickness, temperature and salinity structures, as described by Brown and Irish (1993). In one notable case, the replacement of the colder, fresher Winter Intermediate Water by the warmer, saltier Summer Intermediate Water (with origins in the eastern Gulf) is demonstrated clearly. In another case, a vertical section of the layer distributions during February shows the extension of bottom and intermediate layers from Wilkinson Basin up onto Georges Bank. Thus there is a potentially "easy" pathway by which Calanus finmarchicus can move onto the Bank after overwintering in the bottom of the Wilkinson Basin. Climatological Box Modeling The monthly climatological hydrographic fields are being used in a 48-box model to infer advection and diffusion between the four layers and twelve different regions of the Gulf. An inverse model, which is based on the works of Wunsch and Minster (1982), Bolin et al. (1987) and Fiadeiro and Veronis (1977, 1984), has been developed for the Gulf of Maine / Georges Bank region. At present the model, which is based on the conservation of mass, temperature and salinity, is an underdetermined linear system of 144 equations with 300 unknown advective and diffusive transports. Model coefficients are determined from the box-averaged temperatures and salinities which have been derived from the interpolated AFAP fields. Matrix inversion, necessary to determine the unknown transports, is performed using singular value decomposition (SVD). This approach provides estimated model solution errors which can be compared to OA error fields to diagnose uncertainties. The initial inverse model calculations for September provide estimates of the climatological structure of transport and mixing fields in the Gulf during late summer to fall. We have focused first on this time of the year because the seasonal pycnocline isolates the surface layer from the intermediate and deep layers throughout much of the region. Thus we minimize; (a) anticipated difficulties in specifying surface heat fluxes and net freshwater input and (b) linkages between the surface and deeper layers of the system. The results of the autumn calculations will help us to better understand the sensitivity of model transports to variabilities in boundary flow in a physical system which is somewhat simpler than during the months when vertical exchanges are more vigorous. The next set of model calculations will focus on the winter months, when Calanus finmarchic us is suspected to be overwintering in the bottom of the Wilkinson Basin. For this time period, we will establish surface boundary conditions by developing a climatology of air-sea heat and freshwater fluxes. The more complicated transitional months will then be modeled. A goal of this research is to reduce the indeterminacy and hence the uncertainty in the model solutions. This can be done by adding boundary conditions and/or more tracers, for which can be written conservation statements, to the system. The zooplankton genes studied by Ann Bucklin represent potential tracers and we plan to work with her to construct the appropriate gene conservation statements for the domain of the box model. References: Bolin, Bert, Anders Björkström, Kim Holmön and Berrien Moore (1987) On Inverse Methods for Combining Chemical and Physical Oceanographic Data: A Steady-State Analysis of the Atlantic Ocean, Report CM-71, August 1987, University of Stockholm, Stockholm, Sweden. Carter, E. F., and A. R. Robinson (1987) "Analysis Models for the Estimation of Ocean Fields," Journal of Atmospheric and Oceanic Technology, Vol. 4, No. 1, March 1987, 49-74. Brown, W. S. and Irish, J. D. (1993) "The Annual Variation of Water Mass Structure in the Gulf of Maine: 1986-1987," Journal of Marine Research, 51, 53-107. Bub, F. L., and W. S. Brown (1995) "Seasonal Evolution of the Gulf of Maine Climatological Hydrographic Structure: 1970-1989," EOS 76 (17): S189. Fiadeiro, M. E. and G. Veronis (1977), "On Weighted-Mean Schemes for the Finite-Difference Approximation to the Advection-Diffusion Equation," Tellus 29, 512-522. Fiadeiro, M. E. and G. Veronis (1984), "Obtaining Velocities form Tracer Distributions," Journal of Physical Oceanography, Vol. 14, 1734-1746. Greenberg, D. A. (1983), "Modelling the Mean Barotropic Circulation in the Bay of Fundy and Gulf of Maine," Journal of Physical Oceanography, Vol. 13. Wunsch, Carl and Jean-François Minster (1982), "Methods for Box Models and Ocean Circulation Tracers: Mathematical Programing and Nonlinear Inverse Theory," Journal of Geophysical Research, Vol. 87, No. C8, 5647-5662. Larval Cod and Haddock Trophodynamics and Transport - Further Considerations on the Effects of Turbulence Francisco E. Werner1, R. Gregory Lough2, R. Ian Perry3, Christopher E. Naimie4, Daniel R. Lynch4 and Brian R. MacKenzie5 1University of North Carolina 2National Marine Fisheries Center, Woods Hole 3Department of Fisheries and Oceans, BC, CA 4Dartmouth College 5Danish Institute for Fisheries & Marine Research Using a model-based approach we consider trophodynamic effects on the growth and survival of larval cod and haddock on Georges Bank during late winter/early spring. We examine: (i) larval search behavior and its effect on encounter with prey, (ii) the ability of larvae to pursue and capture prey in a turbulent environment, and (iii) the effect of turbulence on the dispersion of larvae in the vertical. These studies represent an extension of results described in Werner et al. (1995a) wherein the effect of turbulence-enhanced larval-prey contact rates increased the effective prey concentration resulting in growth of cod larvae consistent with observed rates in the field (see Figure 1). In recent studies (Werner et al. 1995b) we have found that search behavior, the effect of turbulence on pursuit and capture, and vertical dispersion, can decrease the predicted larval growth rates. These results suggest that larval feeding behavior, and especially the ability of larvae to pursue encountered prey, could be an important input to larval growth and survival models. Until larval feeding behavior in turbulent environments is better described, interpretations of how turbulence affects ingestion and growth rates in populations of larval cod and haddock should be regarded as preliminary. The inclusion of turbulence in determining the position of passive larvae in the water column allows the larvae to sample the entire water column contributing to a decrease in the variance of the size of the larvae over time. The ability of larvae to swim and aggregate in the vertical will be necessary to reproduce distributions observed in the field. References Werner, F.E., R.I. Perry, R.G. Lough and C.E. Naimie (1995a) Trophodynamic and Advective Influences on Georges Bank Larval Cod and Haddock. Deep Sea Research II, in press. Werner, F.E., R.I. Perry, B.R. MacKenzie, R.G. Lough and C.E. Naimie (1995b) Larval Trophodynamics, Turbulence, and Drift on Georges Bank: A Sensitivity Analysis of Cod and Haddock. ICES Annual Science Conference, 21-26 September 1995, Aalborg, DENMARK, ICES C.M. 1995/Q:26. Figure 1. Post-hatch time history of cod larvae with cruise behavior and Ff=1:1. Top panel, the percentage of larvae alive (solid line), starved on-Bank (solid line with crosses), and advected off the Bank (solid line with open circles); second panel, the daily size distribution (ug) for the live larvae on the Bank, the 15% per day growth curve, the death barrier and the mean daily weight of those live larvae still on the Bank. Also shown are the horizontal distribution of the live larvae (bottom left panel) and their vertical distribution relative to the local bottom-depth (bottom right panel). Modeling Zooplankton Population Dynamics in Realistic Flow Fields Daniel R. Lynch and Wendy Gentleman Dartmouth College As part of the Lynch et al. modeling project, we have developed highly detailed 3-D circulation fields on realistic Georges Bank topography [Lynch et al. 1995; Naimie,1995]. These flow fields have been used in detailed physical-biological studies involving larval fish IBM's (individual-based models) incorporating trophodynamic relationships [Werner et al., 1995]. In those studies, the prey field is specified exogenously, and starvation on-bank is found to be a significant factor for both cod and haddock. Here we present the modeled transport (advection and dispersal) of stage-structured zooplankton populations in these flow fields. Our intentions are: * to develop realistic, dynamic prey fields to support the larval fish studies * to explore the likely source regions for Georges Bank zooplankton * to assist in the interpretation of field data collected during the GLOBEC field years * to develop a computational basis for evaluating the essential population dynamics of the target species. The result is the detailed interaction of tidal-time hydrodynamics with the insitu population dynamics of a principal target species in the NW Atlantic Globec program. The model displayed here is for Pseudocalanus sp., as described by Davis [1984]. We use 13 stages as depicted in Table 1; each stage is represented as a CBM (concentration-based model). All 13 concentrations are transported in the flow, with appropriate interactions. Note that at C, the maturation time from egg to adult is approximately 55 days, which is comparable to the recirculation time on the shallow cap of the Bank in late winter Figure 1). Note also that the survivorship through this maturation period exceeds the adult death rate, such that the population will experience unstable growth under these conditions. In Figure 2 and Figure 3 we display the results of a representative computation with a Gaussian pulse of adult females introduced on the northwest peak. The subsequent development follows qualitatively the expected dynamic. The detailed flow pattern creates a bifurcated population with a portion recirculating around the shoals, and another portion recirculating via the northeast peak. The delayed onset of instability is evident in the early decay of the adult population and its subsequent growth evident at 60 days. References Davis, C. S. 1984. Interaction of a copepod population with the mean circulation on Georges Bank, J. Mar. Res. 42: 573-590 Lynch, D.R., J.T.C. Ip, C.E. Naimie, and F.E. Werner, 1995. Comprehensive coastal circulation model with application to the Gulf of Maine, Continental Shelf Research, in press.Naimie, C.E., 1995. Georges Bank residual circulation during weak and strong stratification periods - Prognostic numerical model results, J. Geophysical Research, in press. Werner,F.E., R.I. Perry, R.G. Lough, C.E. Naimie, 1995. Trophodynamic and advective influences on Georges Bank cod and haddock, Deep Sea Research II, Topical Studies in Oceanography, Special Volume on Georges Bank; Wiebe and Beardlsey, eds. In press. Table 1: Time Constants (days, at °C) by Stage. Figure 1: Illustrating the recirculation structure on the bank. top Figure 2: Female distribution at selected times. Figure 3: Egg distribution at selected times. Tracking the advection of cod and haddock larvae along the southern flank of Georges Bank: Spring 1995 James P. Manning1, Christopher E. Naimie2, R. Greg Lough1 1National Marine Fisheries Service 2Dartmouth College The distribution of gadid larvae was determined from a series of cruises on Georges Bank in the spring of 1995. High concentrations of eggs were found on the northeast peak (66 37'W, 90m) in March, early stage larvae were found on the southern flank (67 22'W, 90m) in April, and older stage larvae were found further west (68W, 65m) in May just inside an encroaching shelf/Slope Water front. (See Figure 1.) Records of velocity from moored and drifting instrumentation on the southern flank document a series of along-bank burst of wind-driven flow (15km/day) as well as periods of sluggish or, in some cases, reversed (eastward) subtidal advection. Extensive CTD surveys depict the foot of the shelf/Slope Water front that encroached as far as the 65 meter isobath. Did these low-frequency (2-5 day) flow events contribute significantly to the resultant advective pathways? Given a 3-d prognostic time stepping model with observations of real wind at a nearby NOAA buoys and climatological density fields, selected periods of advection have been hindcast over several tidal cycles. Near-surface (15 meters) modeled flows are compared with the GPS drifters drogued at 15 meters and Eulerian current observations at 15 meters. The model demonstrates its ability and potential to simulate the advection of cod and haddock from the time and place of spawning through their early life stages. Seasonal variation in cod and haddock egg distributions on Georges Bank in relation to particle residence times Fred H. Page1, Michael Sinclair2, Christopher Naimie3, W. Smith (retired)4, Peter Berrien4, John Loder2, Greg Lough5 1Department of Fisheries and Oceans, CA 2Bedford Institute of Oceanography, CA 3Dartmouth College 4National Marine Fisheries Service, Sandy Hook Laboratory 5National Marine Fisheries Service, Woods Hole The team of modelers headed by Dan Lynch has developed a suite of physical, biological and bio-physical numerical models that have been used to describe and investigate the circulation over Georges Bank and its influence on the distribution and trophic interactions of the cod and haddock early life stages. Descriptions of the physical and trophodynamic models are presented elsewhere in this workshop. In applications to date of the biological model we have released particles in the core spawning area, the northeast peak of Georges Bank. In this presentation we describe the bi-monthly (Jan-Feb, Mar-Apr, ...) horizontal distributions of cod and haddock spawning, as indicated by early stage eggs, over the entire Bank and compare these to model based estimates of the bi-monthly distributions of residence times of particles released over the Bank. The bi-monthly distributions of cod and haddock early stage eggs are estimated from the 1978-1987 MARMAP dataset by compositing over years. As expected the largest numbers of cod and haddock eggs are found in the March-April period. Smaller, although still significant, numbers of cod eggs are found in Jan-Feb, May-Jun, Sept-Oct and Nov-Dec and smaller numbers of haddock eggs are found in Jan-Feb and May-Jun. The highest concentrations of eggs are consistently located over the northeast peak of the Bank. However, the distribution of cod eggs extends over most of the Bank with the exception of the southern flank, whereas haddock egg distributions are more restricted, being mainly found over the eastern portion of the bank and over the Great South Channel. The residence times of water on Georges Bank were estimated for each of the bi-monthly periods by tracking model particles released in the Naimie et al. three dimensional numerical model of the circulation on Georges Bank. The model includes forcings by the tide, bi-monthly mean wind stress, a Scotian Shelf inflow and a diagnostic calculation of the baroclinic flow field. The particles were released at 1, 10, 20, 30, 40 and 50m depth on a 10km square grid placed over the Bank. Each particle was advected by the three dimensional flow field for 60 days. The time when each particle left the bank was noted and the residence times for each depth were averaged to produce a depth averaged residence time for each release site. These times were then contoured to produce maps of the bi-monthly depth averaged residence times (Figure 1). The spatial distribution of the residence times varied seasonally. The zone of longest residence times (60+days) was centered within the 60m isobath. The southern flank of the bank consistently had the shortest residence times and the residence times tended to increase with release depth in most areas. As the season progresses the area of the longest depth averaged residence times expands toward the east and south. In Jan-Feb the longest depth averaged residence times are centered over the northwest portion of the Bank within the 60m isobath and in Jul-Aug the 60d residence time contour encompasses much of the area within the 60m isobath. During March-April, when the cod and haddock eggs are most numerous the depth averaged residence times in the spawning areas are greater than 40 days and in much of the area are greater than 50 days. The residence times at 30m for this area and period are generally greater than 60d. The preliminary indication is therefore that the cod and haddock are spawning in the areas and at times of longest residence time. Figure 1: Composite distributions of cod and haddock early stage eggs (Top left and top right panels) and bi-monthly depth averaged residence times (days) of model particles released over Georges Bank during four bi-monthly periods. Each egg composite represents the combined data from all cruises within the 1978-87 MARMAP dataset. Case Studies of Retentive Circulation Features of Cod-Haddock Eggs and Larvae on Georges Bank in 1978, 1980, 1983, and 1985 Greg Lough1, Peter Berrien1, James Manning1, Francisco Werner2, John Loder3, Fred Page4, Charles Hannah3, Christopher Naimie4, Ian Perry5, Michael Sinclair3, and Daniel Lynch4 1National Marine Fisheries Laboratory 2Univerity of North Carolina 3Bedford Institute of Oceanography, CA 4Department of Fisheries & Oceans, N.B. CA 4Dartmouth College 5Department of Fisheries and Oceans, BC, CA Diagnostic modeling results to date indicate that between year differences in circulation processes can substantially impact retention of eggs and larvae on Georges Bank and contribute to variable recruitment (Lough et al. 1994). In 1982 there was an apparent offshore loss of eggs and early larvae during April, where as in 1985 there was a westward displacement of eggs and larvae from the northeast spawning ground but little evidence of offshore loss. Our goal is to look for common physical factors in the 11-year MARMAP data (1977-87) that may be involved in the production of good and bad year classes of cod and haddock. Good year classes (age 1) of both cod and haddock occurred in 1978, 1980, 1983, 1985, and 1987. From an examination of the MARMAP data, eggs and larvae appeared to be retained on the shoals to a greater degree during these years, with the exception of 1987 which was a year of unusual westward transport, but still high recruitment. Retention on Georges Bank within the 60-70-m isobath is probably due to strong recirculation and few disruptive events. These years of apparently strong retention will be analyzed using a prognostic finite-element model on realistic Georges Bank topography to determine the 3-D residual circulation (Naimie, 1995). A particle tracking model will be used to simulate the drift of eggs and larvae from their spawning sites under realistic monthly climatological scenarios. The model requirements include 1) Scotian Shelf inflow, 2) May-June baroclinic component, 3) advective-diffusive coupling, 4) weekly wind stress, 5) buoyancy specification of egg stages, and 6) vertical migration model of larvae. References: Lough, R. G., W. G. Smith, F. E. Werner, J. W. Loder, F. H. Page, C. G. Hannah, C. E. Naimie, R. I. Perry, M. Sinclair, and D. R. Lynch. 1993. The influence of advective processes on the interannual variability in cod egg and larval distributions on Georges Bank. ICES mar. Sci. Symp. 198: 356-378. Naimie, C. E. 1995. Georges Bank residual circulation during weak and strong stratification periods - Prognostic numerical model results. J. Geophysical Res. (Submitted) A Coupled Physical-Biological Model of Georges Bank Peter J. S. Franks1 and Changsheng Chen2 1 Scripps Institution of Oceanography. 2University of Georgia A coupled physical-biological model of the plankton on Georges Bank was developed. The physical model was a modified version of Blumberg and Mellor's (BM) primitive equation model with Mellor and Yamada level 2.5 turbulent closure scheme (Blumberg, 1994; Chen and Beardsley, 1995a). The biological model was the simple three-component nutrient-phytoplankton-zooplankton (NPZ) model of Franks et al. (1986). The model was initialized with a linearly stratified temperature field representative of summer, and a steady-state solution of the NPZ model (no horizontal dependence). The model was forced by the M2 tide at the southern open boundary. The model results showed that the strong tidal currents vertically mixed the water column on the top of the bank, creating a tidal mixing front at the 40-m isobath on the northern flank and at the 50 to 60-m isobath on the southern flank. Correspondingly, biological fields were homogenized on the bank, while a high concentration of phytoplankton developed at the fronts on both the northern and southern flanks. When a sinking velocity of 1 m/day was taken into account, the model predicted a subsurface maximum of phytoplankton over the slope at about 15-20 m below the surface (Figure 1). The biomass and fluxes of biological variables in the model agreed well with field measurements from Georges Bank. The phytoplankton in the well-mixed region of the bank was found to be nutrient replete with an f ratio of about 0.3. The phytoplankton patches at the fronts were supported by vertical fluxes of nutrients from the slope, below the euphotic zone. Physical studies using Lagrangian trajectories indicated that the up-slope fluxes of nutrients were caused by up-slope residual Lagrangian advection associated with the temporal variation of the thickness of the sloping bottom boundary and the reflection of internal tides. (Figure 1:) Panel Discussion The Life History Paradigm for Calanus finmarchicus Charles B. Miller Oregon State University Our basic understanding of the usual seasonal pattern of life history events in Calanus finmarchicus was provided in a 1934 paper by S. Marshall, A. G. Nicholls and A. P. Orr (1934, JMBAUK, 19: 793-828). Their study considered the population in the Clyde Sea area of western Scotland. Half of the year is spent by the Calanus stock resting at depth in the fifth copepodite (C5) stage. In February this resting stock completes its maturation as generation 'Go', C6 males appearing before females. The females ascend to the upper water column and commence spawning of generation 'G1' when the spring bloom provides copious food to support egg production. G1 requires about 2 months to develop, maturing immediately without rest and producing G2. G2 develops slightly faster, then in July or early August returns as C5 to depth to repeat the cycle. Variants on this basic theme have been noted, many of them reviewed in Marshall and Orr's book (1955, The Biology of a Marine Copepod, Calanus finmarchicus (Gunnerus), Oliver and Boyd, Edinburgh, 180 pp.). For example, fjord populations north of central Norway have strictly one generation per year, C5 of G1 returning to depth. Kaartvedt (pers. comm.) suggests this is because continuous summer illumination provides no time near the surface when visual predators cannot see Calanus. Resting stocks at most locations turn out not to be entirely C5, but to include a fraction of C4. The depths at which C5 rest vary greatly across the range. In New England Slope Water the resting stock centers around 470 m. In the basins of the Gulf of Maine and Scotian shelf it resides just above the basin bottom. In the eastern Atlantic it is at and below 800 m. The Georges Bank program has observed a number of new variants on the standard theme. In November, 1994, Durbin, Miller et al. observed a sizeable fraction of the C5 stock in the upper 30 meters of the water column, actively feeding both day and night. These animals showed no preliminary signs of breaking their rest, such as gonadal or adult tooth development. Probably they found a period in which the surface was not patrolled by visual predators and took advantage of it. Meise has determined from MARMAP samples that a small fraction of the Gulf of Maine stock continues active reproduction and development right through the year. Apparently the resting stage is not obligatory. We have observed in both 1994 and 1995 seasons that some fraction of each spring generation (G1, G2, possibly G3) initiates rest, while another group matures and spawns immediately. We don't know what determines which of these paths a given individual will take. Genetically determined behavior based on timing cues for the season (e.g., photoperiod changes) could vary among individuals, or they could somehow receive different cues from the habitat. We do not, yet, have a method for testing these hypotheses. Major life history issues currently under investigation by us and by European participants in the TransAtlantic Studies of Calanus program include evaluations of (1) the extent to which developing stocks over shoal continental shelves (Georges Bank, the North Sea) are supplied from deep basins or offshore and (2) the extent to which the highly productive shelf populations of spring return resting C5 to deep waters. (3) It is likely that some genetically male Calanus mature as females. Georges Bank and Norwegian fjord stocks are being examined to determine frequency and seasonal dependence of this sex change. (4) Studies throughout the range will determine the dependence of fecundity in Calanus upon seasonal timing, food availability, temperature, and other factors. We are fine tuning our paradigm for the life history of Calanus.](sullivan1b-figure1.gif>Figure 1</a>.)<BR>
<BR>
<p><hr><p>
<CENTER><B> Spatial Changes in the Density Distributions of Larval Cod and Haddock, and their Fish Predators on Georges Bank during the Springs of 1994 and 1995<BR>
<BR>
</B>William L. Michaels, Michael J. Fogarty, Gina Reppucci and Mark Robinson<BR>
<BR>
National Marine Fisheries Service, Woods Hole<BR>
<BR>
</CENTER> <BR>
One objective of the NOAA Coastal Ocean Program's (COP) Georges Bank Predation Study is to understand the impact of predation on larval and early-demersal cod and haddock. Predation is unquestionably one of the critical mechanisms affecting the survival of larval cod and haddock on Georges Bank, particularly when the pelagic fish biomass are presently at record high levels (e.g, Atlantic mackerel and Atlantic herring biomass each exceed 2 million metric tonnes). Historically, populations of mackerel and herring undergo northward spring migrations across the southern flank of Georges Bank about May, coinciding with the drift of larval cod and haddock. We are investigating the hypothesis that temperature-driven fluctuations in predation inflicted by these fish migrations affect cod and haddock recruitment on Georges Bank.<P>
Systematic grid surveys (replicated twice in May 1994 and three times in May 1995) were sampled simultaneously for ichthyoplankton (using 61cm bongo), fish predators (bottom and midwater trawls, and acoustics), and hydrography (e.g., CTD). Spatial changes in the density and distribution of larval cod and haddock on southern Georges Bank were shown in relation to density distributions of fish predators. Preliminary analysis of mackerel and herring diets indicate they fed predominantly on planktonic prey such as amphipods, copepods (including <I>Pseudocalanus</I> and <I>Calanus finmarchicus</I>), decapods, chaetognaths, hydroids, and larval fish. DNA and immuno-assay probes are presently being tested to identify well digested cod and haddock larvae from fish stomachs. Further analysis will continue to examine the hypothesis that temperature-dependent episodic predation contributes to the spatial changes in the density distribution of larval cod and haddock.<P>
Additionally, the 61 cm bongo sampler (and most likely the 1m� and 10m� MOCNESS samplers) did not adequately sample the population of age-0 cod and haddock during May, as shown by the large numbers of post-larval cod and haddock (ranging 18-75mm) captured by the Shuman midwater trawl. For this reason, midwater trawling during May and June is an important consideration with broad-scale sampling to obtain abundance and mortality estimates for cod and haddock needed by the GLOBEC population dynamics initiative. We believe the COP-GLOBEC partnership is highly advantageous for accomplishing the goals of both programs.<P>
<p><hr><p>
<CENTER><BR>
<BR>
<BR>
<B>Storage Lipids in<I> Calanus finmarchicus </I>from Georges Bank and Southern Gulf of Maine</B><BR>
<BR>
Charles B. Miller and C. A. Morgan<BR>
<BR>
Oregon State University<BR>
</CENTER><P>
<I>Calanus finmarchicus,</I> a dominant member of winter-spring plankton in waters surrounding Georges Bank, stores organic matter in the form of lipid. Lipid storage is especially prominent in the fifth copepodite (C5) stage which often undergoes a rest phase from early summer through early winter. When a C5 is fully loaded, about 85-90% of the store is contained in a tubular oil sac running through the core of the prosome. A similar fraction is constituted of wax esters (WE), the remainder is triacylglyceride (TAG). We have studied the lipid storage process in C5 by (1) video imaging of live specimens at sea to measure sac volume and (2) silica rod chromatography with flame ionization<BR>
detection of lipid peaks (Iatroscan)..<P>
Our observations suggested in several ways that all of the oil sac content must be WE with TAG dispersed elsewhere. We tested this by micropipette withdrawal of oil sac contents followed by Iatroscan analysis of both oil and the rest of the body. This partitioning produced dramatic reduction in TAG relative to WE in withdrawn oil when compared to the rest of the body (Figure 1). Thus, oil sac contents must be almost pure WE. This was stated in 1972 by Benson et al., but no data were shown.<P>
In samples from May-June of 1994 we discovered a discrepancy of roughly two-fold between video estimates (greater) of oil sac content and Iatroscan estimates (lesser) of WE. A long search for the source of this problem showed strong variation in Iatroscan response factors (peak area relative to mass of lipid loaded) among slightly different WE. To obtain a calibration curve suitable for lipids from <I>C. finmarchicus</I>, we performed a chromatographic separation of WE and TAG from specimens in bulk ('strawberry daquiri'). A calibration curve for copepod WE (Figure 2) showed response factors about half those of our normal standard (palmitoyl palmitate), fully explaining the video vs. Iatroscan discrepancy. From that point we have used the copepod WE as our calibration standard. Iatroscan data have established many of the hypotheses about lipid storage in the Calanidae, some of which will need to be revised.<P>
Our 1995 data reveal a strong pattern in the relative amounts of TAG and WE in C5 collected on April to July BroadScale cruises (Figure 3). This pattern can be modelled from any one point (e.g., fixing TAG at 17 g estimated from the 34% TAG found at 50 g total storage lipid {TSL}, all other points are well predicted by varying TSL). Clearly animals preparing storage for the rest phase have a TAG quota, above which all additional TSL is as WE. Probably the TAG serves as a transfer form in the body. When it reaches a threshold concentration, WE synthesis is enabled and conversion to WE holds TAG constant. Animals collected on February and March cruises did not follow this pattern (Figure 3), although the curve of the one point model set a lower limit to the fraction of TAG present. Many late winter specimens had much more TAG. Since TAG is the storage lipid deposited in eggs, the C5 of late winter are presumably converting WE back to TAG in preparation for egg production. Recent experimental evidence (L. Fessenden, pers. comm.) with C. pacificus suggests that exactly this transformation occurs.<P>
Video estimates of oil sac volume plotted against prosome length showed for all sampled months (June data are given as an example in Figure 4) that maximum oil sac volume in short animals is disproportionately less than that in long ones. Given approximately constant shape, a length change from 2.0 to 2.5 mm should correspond to a volume increase near two-fold (2.53/1.853 = 2.47). The estimated oil sac volume difference is of order eight-fold. Apparently there is an extreme penalty in oil storage capability at the small end of the size range. We have not explained this, yet.<BR>
<BR>
Figure 1. <I>Calanus finmarchicus</I>. Iatroscan chromatograms for oil sac content withdrawn by micropipette (dashed) and for the remainder of the bodies of 5 C5.<P>
<BR>
<BR>
<BR>
<BR>
<BR>
<BR>
Figure 2. Iatroscan calibration curves for a variety of commercially available waxes (dark line at left for our usual standard, palmitoyl palmitate) and for WE chromatographically purified from bulk <I>Calanus finmarchicus </I>(dark line at right).<P>
<BR>
<BR>
<BR>
<BR>
<BR>
<BR>
Figure 3. <I>Calanus finmarchicus </I>, C5. Fraction of triacylglycerides (TAG) plotted against total storage lipid (TSL) mass estimated by Iatroscan in April, May, June and July samples from the Georges Bank BroadScale Survey. Each point represents a single specimen. Open box is a single point selected as the basis for a ){kind=link}