INVESTIGATORS: Ann Bucklin (PI) Ocean Process Analysis Laboratory University of New Hampshire Durham, NH 03824 acb@kepler.unh.edu Wendell Brown (co-PI) Ocean Process Analysis Laboratory University of New Hampshire Durham, NH 03824 wsb@panther.unh.edu GRANT PERIOD: July 1, 1993 - January 31, 1996
The next phase of the population genetic studies is now underway. The goal of this phase is to investigate smaller-scale patterns of gene flow of C. finmarchicus within the western N. Atlantic. We have selected several new mitochondrial genes for use as markers of sub-regional scale population genetic structure. MtDNA-enriched extractions were used to obtain sequences for portions of four protein-coding, mitochondrial genes. We have also sequenced these regions for additional copepod and euphausiid species and have used the multi-species alignments to design new amplification and sequencing primers.
Using a 700 base pair region of the mitochondrial cytochrome oxidase I gene, we are now engaged in the molecular population genetic analysis of C. finmarchicus from samples collected during the June, 1994, U.S. GLOBEC Georges Bank Study Broad Scale Survey cruise (in which four members of this laboratory participated). We have expanded the sampling domain for 1994, and have obtained zooplankton samples containing numerous C. finmarchicus from the Labrador Current, Gulf of St. Lawrence, and Scotian Shelf. Molecular analysis of individuals collected across this domain in 1994 will be used to describe gene flow patterns within the western N. Atlantic.
During the next year, we will assay microsatellite DNA variation [also called variable number, tandem repeat (VNTR) loci] in Calanus finmarchicus. One possible advantage of VNTR loci as indicators of dispersal is their marked variability within species. Since the sequences do not encode functional gene products; these non-coding regions are frequently highly variable (Jeffreys et al., 1985). Another advantage of VNTR loci as population genetic indicators is their ease of assay. Samples of hundreds of individuals can be assayed rapidly and relatively inexpensively, once the development work has been completed (see Quellar et al., 1993). The technical steps involved in identifying VNTR loci are well documented in the literature (Jeffreys et al., 1985; Quellar et al., 1993). Our task will be to modify the established techniques to allow the analysis of intraspecific variation of copepod populations.
The identification and characterization of VNTR loci for population studies of Calanus finmarchicus will involve designing oligonucleotide primers that span the VNTR loci and assessing which VNTR loci are sufficiently variable to be useful markers of population structure. Once this stage is completed, individual copepods can be assayed for variability at numerous VNTR loci rapidly. Assay of thousands of copepods collected from the Labrador Current, Gulf of St. Lawrence, Scotian Shelf, Gulf of Maine, and Georges Bankis is planned, to examine patterns of gene flow of Calanus finmarchicus in the western N. Atlantic Ocean.
We are presently setting up the time dependent box model based on the conservation of mass, salinity and temperature and constrained by thermal wind relationships. The first model calculations will provide a climatological picture of the structure of transport and mixing in the Gulf for late summer, when the surface layer is virtually isolated from the intermediate and deep layers throughout much of the region. Focussing on this time period will minimize the anticipated difficulties with surface heat fluxes and net freshwater input; vertical and horizontal mixing; and the transports through the open boundaries.The results of this calculation will help us to better understand the vertical mixing coefficients and the sensitivity of model results to boundary flow variabilities.
The next set of model calculations will focus on the winter months, when the copepod, Calanus finmarchicus, enters diapause in the bottom waters of Wikinson Basin. For this time period, we will develop air-sea surface heat flux climatology to keep track of heat and water mass fluxes which are known to be important during winter. If time allows, we will then attempt to do model calculations for the spring months.
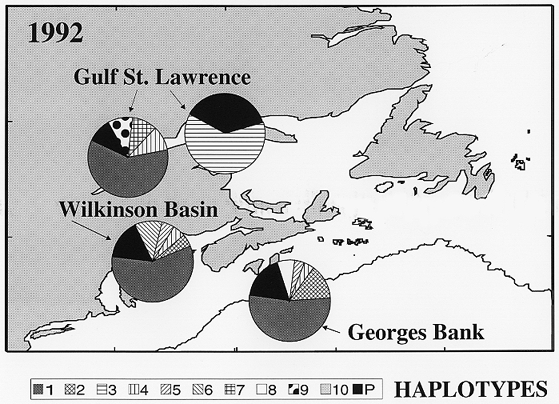
Figure 1. Haplotype abundances by region for 1992. Samples within each region were pooled to yield the regional pattern of haplotype distribution shown. The regions are the estuary of the Gulf of St. Lawrence (STL1), the northern Gulf of St. Lawrence (STL2 and STL3), Georges Bank (GB), and Wilkinson Basin in the Gulf of Maine (WILK).
The only regional differences seen were between pooled samples of the western N. Atlantic and those of the Norwegian Sea (based on data from Bucklin et al., 1995b). There were no significant differences in haplotype frequencies among the samples collected in 1993 and fewer haplotypes were observed in these samples (Figure 2). Intraspecific molecular variation was typical of other marine species: there were 29 haplotypes among the 182 individuals sequenced. The frequency distribution of the haplotypes was highly skewed: 128 individuals shared one haplotype and 19 individuals were unique. There were 24 variable sites among the 350 bases sequenced; estimated nucleotide diversity was 0.0042.
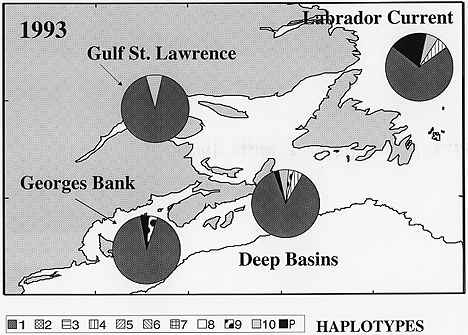
Figure 2. Haplotype abundances by region for 1993. Samples within each area were pooled to reveal regional patterns of haplotype distribution. The regions are the Labrador Current, the Gulf of St. Lawrence (STL1 and STL4), Georges Bank (GB), and the deep basins the Scotian Shelf and the Gulf of Maine (DB).
The genetic character of C. finmarchicus populations in the western N. Atlantic was stable over time in that three of the haplotypes (including the most abundant) occurred in both 1992 and 1993. However, haplotype frequencies differed significantly between the two years. The lack of regional structure in the 1992 samples and the genetic homogeneity of samples collected in 1993 across the domain from the Labrador Current to the Gulf of St. Lawrence to Georges Bank and the Gulf of Maine indicated that there is significant gene flow across this region. The persistent genetic pattern suggests that the Gulf of St. Lawrence may be an important source region for recruitment of C. finmarchicus to Georges Bank. Ongoing studies are underway to examine additional molecular traits (both nuclear and mitochondrial) and to use additional analyses to infer patterns and pathways of dispersal within the western N. Atlantic domain. Determination of zooplankton dispersal patterns within high gene flow species will provide information that may not be determined by conventional oceanographic analyses.
In a related study, the base sequence of a 220 base-pair region of 16S rDNA was determined for six abundant copepod species on Georges Bank (Calanus finmarchicus, Centropages typicus, C. hamatus, Temora longicornis, Pseudocalanus moultoni, and P. newmani) and six species from adjacent N. Atlantic regions (Calanus glacialis, C. helgolandicus, C. hyperboreus, Metridia lucens, M. longa, and Nannocalanus minor). Typical levels of sequence difference were dependent upon taxonomic separation: 5 - 10% between sibling species, 15 - 20% between non-sibling congeneric species, and >30% between species in different genera or families. The phylogenetic reconstruction resolved congeneric and confamilial species groups, and was consistent with the morphological phylogenies of these groups. In addition, the molecular analysis supported the description of sibling species of Pseudocalanus (Frost, 1989) and divided one species, Nannocalanus minor, into three genetically distinctive groups that probably constitute different species. This study demonstrated that the base sequence variation of mitochondrial 16S rDNA is useful for phylogenetic analyses from the subspecific to the familial level. Rapid molecularly-based identification (by sequence-specific oligonucleotide probes and/or restriction enzyme digestions) will be possible based on 16S rDNA sequence and will enable oceanographers to discriminate species of copepods at all life stages.
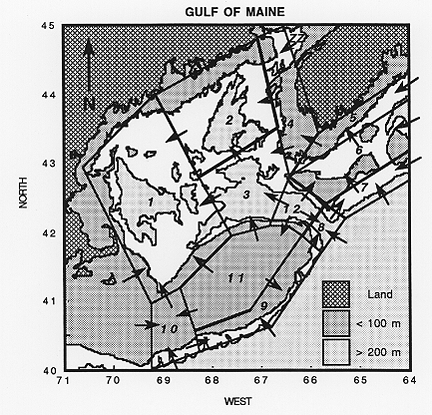
Figure 3. Division fo the Gulf of Maine region in twelve subregions based on Maine water masses. The subregions are the basis of the hydrographic analysis and box modeling.
As winter progresses, all layers cool and continue to freshen. The least dense surface layer all but disappears by February, replaced at the surface by the expanding intermediate density layer (winter layer). Patterns suggest thickening cells of the intermediate layer in Wilkinson basin. Towards winter's end, the climatological analyses show an isopycnal pathway within bottom water, whereby Calanus finmarchicus can move between the over- wintering site in Wilkinson basin and the spring resurgence on Georges Bank (Figure 4). During spring, warming and salting occur as the surface layer expands and summer stratification sets up. Surface warming is first evident in the Georges Bank and then Wilkinson Basin subregions.
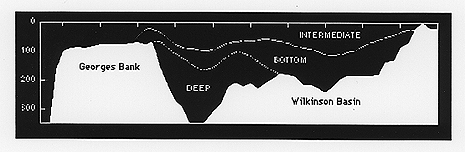
Figure 4. Cross-section through Wilkinson Basin, showing water mass structure, and an isopycnal pathway within bottom water, by which Calanus finmarchicus may move between the Basin and the Bank.
The seasonal evolution of the volume-averaged hydrography in the different subregions show patterns of warming and salting during summer. The region warms most over Georges Bank. The increasingly stratified surface layer, which expands uniformly to a thickness of 30 m across the Gulf, is isolated from the thinning intermediate and bottom layers. During the fall transition, the surface layer cools and freshens as patterns suggest the westward spread of low salinity water the Scotian Shelf.
Intraspecific mtDNA variation of Calanus pacificus: evidence for subspecific differentiation and population structure of C. p. californicus. Invited speaker at a special symposium, Genetics of California Current Organisms, CalCOFI Conference at Lake Arrowhead, CA (November 1-4, 1993).
Population genetics of Calanus finmarchicus in the North Atlantic. Invited presentation at the ICES Workshop for a TransAtlantic Study of Calanus finmarchicus, Oslo, Norway (April 6-8, 1994).
Gene flow in marine planktonic copepods: what mtDNA can and cannot tell us. Invited speaker and co-chair of a special symposium, Molecular approaches in marine and freshwater biology, ASLO/PSA meeting, Miami, FL (June 5-10, 1994).
Mitochondrial DNA indicators of genetic diversity and structure in marine zooplankton. International Marine Biotechnology Conference, Tromso, Norway (August 7-12, 1994).
Bucklin, A. 1994. Molecular population genetics of the copepod, Calanus finmarchicus, evidence of gene flow between populations in the Gulf of St. Lawrence and Georges Bank. EOS 75:56.
Bucklin, A. 1994. Gene flow in marine copepods: what mtDNA can and cannot tell us. ASLO Abstracts a-12.
Caudill, C. and A. Bucklin. 1994. MtDNA sequence divergence within the copepod, Acartia tonsa, among estuaries of the eastern U.S. ASLO Abstracts a-14.
Bucklin, A., B.W. Frost, and T.D. Kocher. 1995a. Molecular systematics of seven species of Calanus and three species of Metridia (Crustacea: Copepoda). Mar. Biol. 121:655-664.
Bucklin, A. 1995. Molecular indicators of zooplankton dispersal in the ocean. Report to the Internat. Union Geol. Geophys., Amer. Geophys. Union, Washington, DC. (In press)
Bucklin, A. and T.D. Kocher. 1995. Source regions for recruitment of Calanus finmarchicus to Georges Bank: evidence from molecular population genetic analysis of mtDNA. Deep-Sea Res. (In press)
Bucklin, A., R. Sundt, and G. Dahle. 1995b. Population genetics of Calanus finmarchicus in the North Atlantic. Proceedings of the TransAtlantic Calanus Workshop, Oslo, Norway. Ophelia. (In press)
Bucklin, A., E.P. Curry, and N.J. Copley. 1995c. Molecular phylogeny and species identification of planktonic calanoid copepods of Georges Bank based on 16S rRNA gene sequences. Mar. Biol. (In review)