INVESTIGATORS: Dian J. Gifford University of Rhode Island Michael E. Sieracki Bigelow Laboratory for Ocean Science
Omnivory in copepod diet. Planktonic protozoa function as important trophic intermediaries in pelagic food webs by repackaging small bacterial and algal cells into food items which are accessible to larger consumers. Although it has long been obsberved that calanoid copepods which feed in the euphotic zone tend to be omnivorous, they are often treated as if they are exclusively herbivorous in both mathematical models and manipulative experiments. Studies with a number of species in a diversity of marine environments indicate that calanoid copepods consume nano- and microzooplankton in laboratory experiments done under controlled conditions and in the field when presented with natural assemblages of prey. Further, consumption of protozoan prey affects copepod condition and, ultimately, production (reviewed in Gifford 1991).
Copepod feeding experiments. Manipulative experiments were done using natural prey assemblages in which consumption of phytoplankton and protozoa was measured directly by monitoring the disappearance of prey. Feeding was measured at each time series station under prey conditions representative of the mixed layer and/or the pycnocline, as appropriate. Ingestion rates of consumers were measured directly in shipboard experiments using natural prey assemblages. Experiments were done with all life history stages of Calanus sufficiently abundant to be sorted: stages C1-C6 females. The detailed protocol for these experiments is described in Gifford (1993a; b). In brief, the design onsists of 2 treatments, an experimental treatment consisting of the nano- and microplankton prey assemblage incubated in bottles with copepods added and a control treatment in which bottles contain only the prey assemblage. Treatment bottles are incubated for 24 hours on-deck in a water-cooled incubator equipped with a rotating wheel. Samples for size-fractionated chlorophyll, nanoplankton and microplankton are collected at the beginning and end of an experiment and analyzed as described above. Per capita and carbon-specific rates of copepod clearance and ingestion are calculated according to Frost (1972) and Marin, et al. (1986).
Table 1. GLOBEC Georges Bank process cruises 1994-1995.
Cruise Research Vessel Date
CI9407 Columbus Iselin 25 May-16 June 1994
EN259 Endeavor 9-21 January 1995
EN262 Endeavor 23 February-10 March 1995
EN264 Endeavor 27 March-8 April 1995
EN266 Endeavor 26 April-9 May 1995
EN267B Endeavor 7-21 June 1995
Table 2. Experiments with Calanus finmarchicus during cruise CI9407.
Experiment Number Date Copepodid Stage
Cal-1 05-29-94 C6 Female
Cal-2 05-29-94 C5
Cal-3 05-29-94 C4
Cal-4 05-31-94 C6 Female
Cal-5 05-31-94 C5
Cal-6 05-31-94 C5
Cal-7 06-07-94 C6 Female
Cal-8 06-07-94 C5
Cal-9 06-10-94 C6 Female
Cal-10 06-10-94 C6 Female
Cal-11 06-11-94 C5
Cal-12 06-11-94 C5
The prey field. Total chlorophyll-a was generally less than 2ug/L at both the STRATIFIED and MIXED sites. At least half of the chlorophyll standing stock was <5 um in size in all experiments. The nanoplankton was dominated by cyanobacteria and heterotrophic flagellates. The microphytoplankton consisted primarily of large centric diatoms. The microzooplankton assemblage was dominated by aloricate choreotrich ciliates >5 um. Heterotrophic dinoflagellates of the genus Protoperidinium were abundant, as was the autOtrophic ciliate Mesodinium rubra.
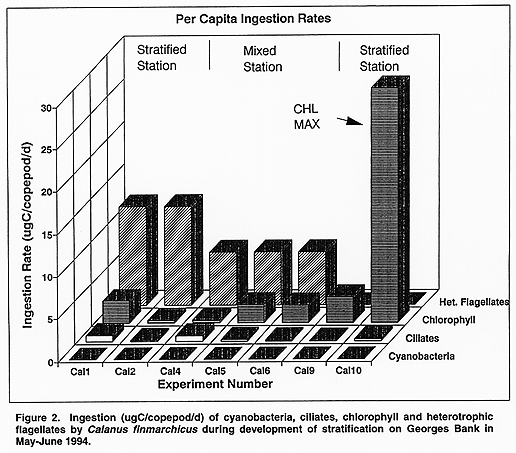
Feeding rates. Ingestion of chlorophyll, cyanobacteria, heterotrophic flagellates and ciliates from a subset of the experiments is shown in Figure 2. As expected, feeding effort was concentrated on particles >5 um, and cyanobacteria were not consumed. Clearance rates (not shown) of ciliates and heterotrophic flagellates greatly exceeded clearance of bulk chlorophyll except in Experiment 10, where the prey assemblage was collected from the chlorophyll maximum. The majority of carbon ingested came from phytoplankton and heterotrophic flagellate prey.
Conover, R.J. 1982. Interrelatuons between microzooplankton and other plankton organisms. Ann. Inst. Oceanogr. Paris 59S: 31-46.
Frost, B.W. 1972. Effects of size and concentration of food on the feeding behavior of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 17: 805-815.
Gifford, D.J. 1991. The protozoan-metazoan link in pelagic ecosystems. J. Protozool. 38: 81-86.
Gifford, D.J. 1993a. Protozoa in the diets of Neocalanus spp. in the oceanic subarctic Pacific Ocean. Progr. Oceanogr. 32: 223-237.
Gifford, D.J. 1993b. Consumption of planktonic protozoa by suspension feeding copepods. pp. 723-727 in: P.F. Kemp, et al. (eds.), Handbook of Methods in Aquatic Microbial Ecology. Lewis Publishers, Boca Raton, FL.
Marin, V. et al. 1986. Measuring feeding rates of pelagic herbivores: analysis of experimental design and methods. Mar. Biol. 93: 49-58.
Sieburth, J. McN. et al. 1978. Pelagic ecosystem structure: heterotrophic compartments of the plankton and their relationship to plankton size fraction. Limnol. Oceanogr. 23: 1256-1263.
Sieracki, M.E. and C. L. Viles 1990. Color image-analyzed fluorescence microscopy: a new tool for marine microbial ecology. Oceanography: 3: 30-36.
[NOT AVAILABLE YET] Figure 1. Drifter station location during May and June 1994. Stations 21, 41 and 43 are the STRATIFIED SITE. Stations 22 and 42 are the MIXED SITE.
[NOT AVAILABLE YET] Figure 2. Ingestion (ugC/copepod/d) of cyanobacteria, ciliates, chlorophyll and heterotrophic flagellates by Calanus finmarchicus during development of stratification on Georges Bank in May-June 1994.