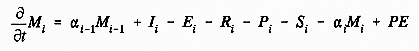
where Mi is the biomass of stage i, alpha-i is the recruitment rate out of stage i, I is ingestion, E is egestion, R is respiration, P is predation, S is starvation, and PE is physical exchange (includes advection and diffusion). For adult and egg life stages, the equation must also include loss and input terms, respectively, for egg production (Figure 3). The left-hand side of the equation can be obtained directly from field data on abundance, and the size distribution and/or age structure of individuals within a population. On the right-hand side, individual growth rate can be measured in lieu of ingestion, egestion, and respiration (Figure 3). Measurements of growth rate (and/or development times) also can provide indices of starvation potential, as well as estimates of transfer rates (i.e., recruitment rates) between life stages (Figure 3). For adult female holozooplankton, egg production rates should be measured as functions of food availability, condition factors, and spawning behavior. Measurements of body size may be important for some holoplanktonic taxa where fecundity is related to adult size. If growth rate and starvation are found to significantly limit population growth, the components of growth can be examined in further detail.
Given that phytoplankton abundance (and proably phytoplankton species composition as well) differs greatly in well-mixed vs. stratified regions of the Bank (shown later in this document in Figure 10), copepod egg production, development and growth rates also may differ between well-mixed, frontal and stratified regions of the Bank. Thus, studies that focus on spatial variations in growth rates could lead to a better understanding of environmental control of copepod population dynamics. Physical process that control phytoplankton dynamics should be studied because whichever physical processes directly control phytoplankton growth rates may indirectly control copepod population dynamics.
Swimming and feeding behavior of the target species could be studied in relation to prey abundance, nutritional quality, and avoidance behavior as well as in relation to microscale patchiness, turbulence, and predation rates (Figure 3). These behavioral activities affect rates of ingestion and respiration (and therefore growth and starvation) as well as susceptibility to predation. Study of these behaviors could provide one of the keys to understanding the relationship between physical processes and population response.
Mortality rates can be estimated from field studies which show abundance of a given developmental stage along with measurements of stage-specific development times, following Steele and Mullin (1977) and Kimmerer (1987). Mortality rates for eggs can be derived from equations in Kiżrboe et al., (1988). Also required will be estimates of physical exchange of animals on and off the Bank from detailed information on the flow field, spatial gradients in animal abundance, and estimates of the time rate of change in abundance. Mortality rates can also be estimated from predator abundances and consumption rates, and prey abundance and prey growth rates.
Physical factors which may directly affect animal abundance include temperature effects on biological rates (Figure 3), microturbulence effects on ingestion, respiration, and predation, and advective losses or gains from the study area. This latter category includes both horizontal advection by mean and episodic currents associated with steady forcing (e.g., buoyancy and tidal) and transient forcing (e.g., storms and mesoscale eddies and rings), and the effective horizontal movement of zooplankton caused by their vertical swimming behavior in a time-dependent vertically sheared current field. Given the apparent long residence time of animals on the Bank, we hypothesize that individuals of some species have evolved behavior patterns which serve to retain them on the Bank. We know little about these behaviors. Also included here is consideration of sources of animals found on the Bank; although larval fish are probably produced locally on the Bank during winter and spring, copepods at that time of the year almost certainly recruit from somewhere else.
At present, we have only a limited understanding of how physical processes interact with the dynamics of dominant zooplankton species on Georges Bank. Local physical processes are of course important, however the effects of large-scale (and remote) physical forcing on ecological efficiency and the consequences for recruitment at higher trophic levels are poorly understood. Significant insight can be gained by modeling interactions between physical transport, mixing-stratification, simple food chain interactions and the population dynamics of dominant or characteristic zooplankton species. Each species has evolved certain characteristics which are affected differently by advective transport out of favorable growth areas; some species may be highly dependent upon seasonal input onto the Bank of individuals from source populations off the Bank, and some may not.